the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 10 Dec 2019
| 10 Dec 2019
Base cations in the soil bank: non-exchangeable pools may sustain centuries of net loss to forestry and leaching
Nicholas P. Rosenstock
Johan Stendahl
Gregory van der Heijden
Lars Lundin
Eric McGivney
Kevin Bishop
Stefan Löfgren
Accurately quantifying soil base cation pool sizes is essential to interpreting the sustainability of forest harvests from element mass-balance studies. The soil-exchangeable pool is classically viewed as the bank of “available” base cations in the soil, withdrawn upon by plant uptake and leaching and refilled by litter decomposition, atmospheric deposition and mineral weathering. The operational definition of this soil bank as the exchangeable (salt-extractable) pools ignores the potential role of “other” soil nutrient pools, including microbial biomass, clay interlayer absorbed elements, and calcium oxalate. These pools can be large relative to “exchangeable” pools. Thus neglecting these other pools in studies examining the sustainability of biomass extractions, or need for nutrient return, limits our ability to gauge the threat or risk of unsustainable biomass removals. We examine a set of chemical extraction data from a mature Norway spruce forest in central Sweden and compare this dataset to ecosystem flux data gathered from the site in previous research. The 0.2 M HCl extraction released large pools of Ca, K, Mg, and Na, considerably larger than the exchangeable pools. Where net losses of base cations are predicted from biomass harvest, exchangeable pools may not be sufficient to support more than a single 65-year forest rotation, but acid-extractable pools are sufficient to support many rotations of net-ecosystem losses. We examine elemental ratios, soil clay and carbon contents, and pool depth trends to identify the likely origin of the HCl-extractable pool. No single candidate compound class emerges, as very strongly supported by the data, as being the major constituent of the HCl-extractable fraction. A combination of microbial biomass, fine grain, potentially shielded, easily weatherable minerals, and non-structural clay interlayer bound potassium may explain the size and distribution of the acid-extractable base cation pool. Sequential extraction techniques and isotope-exchange measurements should be further developed and, if possible, complemented with spectroscopic techniques to illuminate the identity of and flux rates through these important, and commonly overlooked, nutrient pools.
- Article
(1087 KB) - Full-text XML
-
Supplement
(139 KB) - BibTeX
- EndNote





1.1 Importance of soil base cation pools for sustainable land use
In an attempt to decrease net CO2 emissions and to meet the increasing demand for woody biomass products and energy, some forest policies encourage the intensification of biomass extraction from forest ecosystems (Royo et al., 2012; Lauri et al., 2014; Kazagic et al., 2016). These practices may severely impact soil fertility because biomass extraction removes the nutrients elements in the biomass, especially in forest ecosystems where nutrient availability may be low (Feller, 2005; Kreutzweiser et al., 2008; Thiffault et al., 2011; Achat et al., 2015). For example, in the USA it has been projected that increased biomass harvest for biofuels will markedly increase the area of managed forest that will experience net nutrient losses due to biomass export: from 17 % to 50 % of the total forest area for Ca, 20 % to 57 % for K, and 16 % to 45 % for Mg (de Oliveira Garcia et al., 2018). Similar concerns have been raised in other countries, including Sweden (Akselsson et al., 2007a), Belgium (Vangansbeke et al., 2015), Finland (Aherne et al., 2012), and Germany (Knust et al., 2016). Quantifying sustainable levels of biomass extraction, such that soils are not progressively impoverished and productivity can be maintained without nutrient inputs, is therefore a major concern for forest managers and policy makers (Lucas et al., 2014; Achat et al., 2015). The pressures on plant-available base cations result not only from increased harvest intensity but also from continued atmospheric nitrogen and sulfur deposition (Iwald et al., 2013; McGivney et al., 2019), and elevated atmospheric concentrations of CO2 (Duval et al., 2012; Terrer et al., 2016).
The nutrient mass-balance (i.e., input–output budget) approach is commonly used to estimate the net gain or loss of nutrient elements in the soil or ecosystems under different management and climate scenarios (Akselsson et al., 2007a; Knust et al., 2016; de Oliveira Garcia et al., 2018). This mass-balance approach typically accounts for inputs via atmospheric deposition and the weathering of primary minerals as well as secondary clay minerals, and exports occur via leaching and biomass removals (Nilsson et al., 1982; Van Breemen et al., 1984). Under this definition, the mass-balance equation estimates the change in nutrient content in a “source and sink” reservoir of the soil. For base cations, this source and sink reservoir is assumed to be the pool of Mg, Ca, Na, and K stored in the soil as exchangeable cations adsorbed on the cation-exchange complex. This quantity is conventionally measured by ion-exchange soil extractions using concentrated salts (salt-extractable exchangeable pools; Akselsson et al., 2007a; Vangansbeke et al., 2015; Knust et al., 2016).
Accurate estimates of soil nutrient stocks are important for policies on sustainable levels of biomass extraction levels or critical loads, i.e., the maximum level of atmospheric pollutant deposition that will not damage sensitive aspects of the ecosystem, as well as for determining the maximum level of forest biomass harvest, the “critical biomass harvest” that will not result in unacceptable soil and stream acidification (Akselsson and Belyazid, 2018). The net balance of nutrients between influx and efflux in an ecosystem or soil can only be meaningfully interpreted in the context of the magnitude of soil nutrient reserves. If, for example, whole-tree harvesting results in a net decrease of soil Ca, it is equally important to know if this loss is likely to result in significant reductions in the availability of Ca to plants in 1, 10 or 100 forest rotations, as it is to know whether that balance is positive or negative.
1.2 Use of salt-extractable pools in ecosystem mass-balance studies
Base cation nutrient reserves in soil are commonly estimated by measuring salt-extractable quantities, commonly with barium salts or ammonium salts as extractants, to represent what is available to plants (Skinner et al., 2001; McLaughlin and Philips, 2006; Brandtberg and Olsson, 2012; Zetterberg et al., 2016). However, on timescales of years to decades, salt-extractable base cation reserves in the soil do not appear to accurately predict what is available for leaching or biological uptake (Mengel and Rahmatullah, 1994; Bailey et al., 2003; Hamburg et al., 2003; Lucash et al., 2012). This may be due to the presence of other significant pools of base cations in the soil, which are not salt extractable but which contribute to the plant-available pools of base cations. Calcium is known to form strong complexes with organic compounds such as the oxalate ion, which are poorly soluble and not salt extractable (Dauer and Perakis, 2014), but these complexes may be available to plants and microbes over relatively short timescales. The microbial biomass of the soil is quite large, has considerably higher nutrient concentrations than bulk organic matter, turns over very rapidly, and may thus represent a significant pool of available base cations that is missed in salt extractions (Yamashita et al., 2014; Lorenz et al., 2010; van der Heijden et al., 2014). Aluminum and iron oxides and hydroxides may also directly or indirectly (via organic matter or phosphate bridges) adsorb base cations (Kinniburgh et al., 1976; Grove et al., 1981). For example, in a highly weathered tropical soil, Hall and Huang (2017) showed that a significant amount of Mg, Ca and K were sequestered in Fe (hydr)oxide secondary mineral phases, and the microbial dissolution of these phases increased the concentrations of weak-acid-extractable cations. Significant stocks of soil potassium may be stored in the interlayers of clay minerals (Sparks, 1987). This pool of interlayer K is not accounted for by salt-extractable exchangeable cations but may be quite significant for plant nutrition: Falk Øgaard and Krogstad (2005) showed that interlayer K accounted for 26 % to 43 % of K uptake in grassland ecosystems. Not accounting for these additional pools of base cations in the soil may explain why mass-balance models often fail to reproduce the empirically measured change in salt-extractable exchangeable pools (Löfgren et al., 2017; van der Heijden et al., 2014).
Measurements of salt-extractable base cations may be complemented by soil extractions using strong acids. Aqua regia, HF, or lithium metaborate fusion extract the total or near total reserves of elements, which may then be used to (i) estimate the relative distribution of minerals in the soil (Posch and Kurz, 2007) and (ii) estimate the mineral weathering flux based on assumptions of weathering kinetics derived from laboratory dissolution experiments (Warfvinge and Sverdrup, 1992). Moderately concentrated (defined here as 0.1 M–1 M) strong acid extractions (primarily HCl, but also HNO3) have been used to extract nutrient elements from soil organic matter (e.g., Ca from Ca oxalate; Dauer and Perakis, 2014), secondary minerals (e.g., K from clay interlayers; Simonsson et al., 2016) and relatively labile or easily weathered primary minerals (e.g., Ca from apatite, Mg from biotite; Blum et al., 2002; Lucash et al., 2012). However, these extractions are non-specific and will dissolve a range of secondary minerals, likely releasing microbial nutrient pools and partially dissolving non-target granite-derived tectosilicate minerals.
1.3 Study objective
In order to examine the potential importance of non-exchangeable sources of base cations in relation to net losses of base cations from forest harvesting we utilized published data from a particularly well-studied forest site at Kindla, Sweden, to form a mass-balance budget of base cations under different harvest scenarios, and we compared annual losses, where predicted, to base cation pools in the soil as defined by different extractants. The potential for stem-only or whole-tree harvesting to result in net losses of base cations was examined, then these estimated losses were compared to extraction-defined (distilled water, BaCl2 0.1 M, HCl 0.2 M, HCl 0.5 M, and aqua regia) base cation pools to estimate the number of harvest rotations of net nutrient losses these pools could potentially buffer against. The aim of this paper is to discuss (i) what additional pools of base cations in the soil may actively contribute to the “source and sink” pool, (ii) how important in terms of size these additional pools can be, and (iii) how these additional pools may contribute to replenishing plant-available pools over time and under different management scenarios.
2.1 Study area
Kindla is a catchment within the Swedish national integrated monitoring program (Löfgren et al., 2011) and a core site in Long-Term Ecosystem Research (LTER) Europe (https://deims.org/search/all/Kindla, last access: 1 August 2019). The monitoring was initiated in 1995 and is related to the Convention on Long-Range Transboundary Air Pollution – LRTAP 1979 (UNECE) and the International Cooperative Programme on Integrated Monitoring of Air Pollution Effects on Ecosystems (ICP-IM, https://www.unece.org/env/lrtap/workinggroups/wge/im.html, last access: 1 August 2019). It focuses specifically on the monitoring of physical, chemical, and biological processes in time and space to increase the knowledge on causes of ecosystem changes (Starr, 2011). Data from the Kindla catchment are used for biogeochemical process evaluations (see e.g., articles in Starr, 2011), status and trend assessments (e.g., Vuoremaa et al., 2018), and model exercises (e.g., Zetterberg et al., 2014; McGivney et al., 2019). The Kindla site is also designated as one Critical Zone Observatory, and in this article we evaluate data from Kindla soils collected and analyzed within the EU Fp7 project Soil Transformations in European Catchments (SoilTrEC, Banwart et al., 2011). The study was initiated due to lack of forest harvest induced depletion of the exchange Ca and Mg pools even though mass-balance calculations indicated such declines should occur (Olsson and Lundin, 1990).
The forest examined at Kindla is a naturally regenerated, uneven-aged Norway spruce (Picea abies, L.) forest, which has been minimally affected by forestry activities during the last century. Granitic bedrock is overlain by till soils of the same geological origin. Soils from three proximate forest plots were sampled from the Kindla catchment (59∘05′ N 12∘01′ E; Löfgren et al., 2011). The three plots cover an elevation gradient from 300–320 m a.s.l. over a distance of ∼150 m. The upper and intermediate sites are in groundwater recharge areas on Podzols and Regosols, respectively. The lowermost site is located on a Regosol in a groundwater discharge area with considerably more organic matter in the upper mineral horizons (see Supplement Methods and Table 1 for more detailed description).
Table 1Sample description for nine samples from three plots, three depths at each plot, along the hydrology gradient at Kindla.

1 Clay < 0.002; silt > 0.002, < 0.02; sand > 0.02, < 2 mm. 2 Bulk density was estimated for each horizon with the pedo-transfer function from Nilsson and Lundin (2006) based on depth and organic carbon content. 3 Site index according to Zetterberg et al. (2014).
2.2 Base cation fluxes and forest mass balance
For each plot and harvest scenario the mass balance of each base cation, Ca, K, Mg, and Na, was calculated as follows:
where BCdep represented inputs via atmospheric deposition (wet and dry), BCweath represented inputs to the soil via mineral weathering, BCleach represented losses via leaching below the B horizon, and BCuptake represented uptake of base cations into vegetation and subsequent removal at harvest; all terms were calculated per square meter per year. Total deposition (wet + dry) of base cations and mineral weathering rates were taken from McGivney et al. (2019). Base cation uptake into tree biomass and removal via either stem-only (stems + bark) or whole-tree (roots + stump + stem + bark + branches-logging losses) harvest were taken from Zetterberg et al. (2014), McGivney et al. (2019), and Nilsson et al. (2018). Leaching estimates were based on soil water sampled with suction lysimeters (P80 ceramic cups) in the B horizon and compared to catchment runoff (Löfgren et al., 2011). Mineral weathering inputs were taken from modeled values from McGivney et al. (2019). Deposition, weathering, and biomass uptake fluxes were calculated for the entire catchment including the three soil sampling sites located along a hydrological gradient (recharge, intermediate, and discharge areas), while leaching losses were calculated for each of these three hydrological locations (for a more detailed description on how each flux was calculated see Supplement Methods with considerably more organic matter.).
2.3 Soil base cation pools
As part of the EU FP7 funded SoilTrEC project (Banwart et al., 2011) a series of non-sequential chemical extraction were performed on nine composite mineral soil samples, representing the E, B, and C horizons at the three soil sampling sites. Extractions were performed with distilled water, 0.1 M BaCl2, 0.01 M ammonium lactate, 0.2 M HCl, 0.5 M HCl, and aqua regia (3HCl:HNO3). These extractions were filtered and measured for elemental contents of base cations Ca, Mg, K, and Na as well as P, Fe, and Mn. Soil pH, carbon, and nitrogen concentrations as well as soil texture and moisture content were also measured for each sample (see Table S1 in Supplement and Supplement Methods for more detailed description). The O horizon was not included in the SoilTrEc project, but the organic horizon of the same hillslope was sampled in 2006 within the EU Life+ project BioSoil (Cools and de Vos, 2010) and extracted with 0.1 M BaCl2, and aqua regia. The SoilTrEC and BioSoil soil sampling pits are not identical, but forest age, structure and management history, hydrologic gradient, soil types, humus layer thickness, and bulk density are identical. Hence, we have used the BioSoil extraction data for estimating these two base cations pools in the organic horizon.
For the soils, elements, and harvest scenarios that have a net elemental loss over time, we examined the potential for different soil base cation pools to sustain those losses. We assumed that ecosystem fluxes (deposition, weathering, leaching, biomass uptake) would remain constant, and calculated the number of 65-year harvest rotations that each extractant-defined base cation pool could offset net ecosystem losses for each harvest scenario–element-soil combination under which a net ecosystem loss was predicted.
3.1 Soil base cation pools
In the mineral soil, concentrations of Ca and Mg in H2O extractions were generally far less (< 1∕10) than those in the BaCl2 extractions, while H2O extractions of K were only slightly less, and H2O extractions of Na were slightly greater than BaCl2 extractions (Fig. 1). Ammonium lactate extractions of all base cations were similar to BaCl2 extractions; BaCl2 extracted nearly the same or slightly more of the bivalent Ca and Mg, while ammonium lactate tended to extract more monovalent K and Na (100 %–150 %). BaCl2-exchangeable pools of Ca, K, Mg, and Na were particularly high in the E horizon of the downslope Regosol in the groundwater discharge area with significantly higher organic carbon content. Within each soil, BaCl2-exchangeable cations tended to decrease with increasing depth. The extracted amounts of Ca, K, and Mg in the 0.2 M HCl extraction were considerably (5–100 fold) greater than the amounts obtained from BaCl2 extraction. As compared to the 0.2 M HCl extraction, 0.5 M HCl extraction yielded approximately 2-fold greater release of Ca, but only about 1.3-fold greater release of K, Mg, and Na. HCl extraction yields of Ca and Mg increased significantly with depth, while HCl extractions of K did not display a depth trend, and on the upslope and midslope soils E-horizon yields of K were higher than K yields from the B horizons. HCl extraction yields of Na exhibited a slight decreasing trend with soil depth. Compared with the 0.5 M HCl extraction yields, aqua regia extracts of Mg and K were approximately 2-fold greater and aqua regia extracts of Ca approximately 6-fold greater, while aqua-regia-extraction yields of Na were slightly lower (3 %) than from 0.5 M HCl. Aqua regia extractions of all base cations increased with depth; this depth trend was strongest for Ca and Mg, weaker for K, and almost absent for Na.
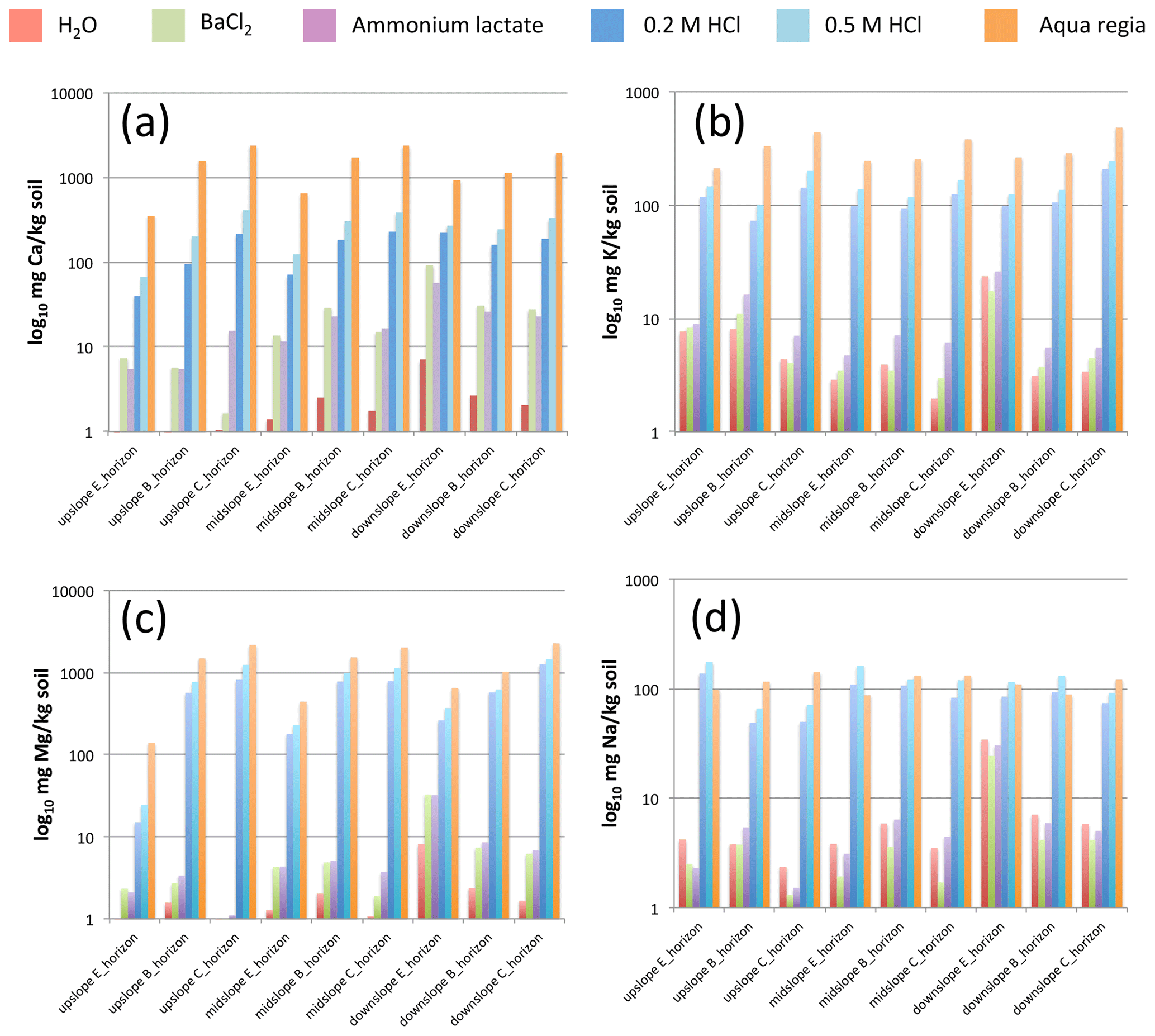
Figure 1Extractable contents of calcium (a), potassium (b), magnesium (c), and sodium (d) across three soils and three soil horizons obtained form six different chemical extractions. Note the log10 scale on y axis.
Humus layer 0.1 M BaCl2-extractable pools of Ca, K, Mg, and Na were 88 %, 100 %, 86 %, and 99 %, respectively, as large as aqua-regia-extracted pools, indicating that nearly all of the humus-associated base cation pools are exchangeable. Humus layer 0.1 M BaCl2-extractable pools of Ca, K, Mg, and Na were 2, 5, 4, and 0.6 times as large as the total mineral soil 0.1 M BaCl2-extractable pools, indicating a predominant role of the humus layer as a reservoir of salt-extractable base cations. Considering only the mineral soil sampled in this study, the BaCl2-extractable pools of Ca. K, Mg, and Na were 12 %, 5 %, 1 %, and 6 %, respectively, of the size of the 0.2 M HCl pools. If we include the humus layer base cation pools, the BaCl2-extractable pools of Ca, K, Mg, and Na then increase to 20 %, 24 %, 3 %, and 9 %, respectively, of the size of the 0.2 M HCl-extractable pools. Hence, 3 %–24 % of the base cations are found in the salt-extractable BaCl2 pool, while the remaining 76 %–97 % are found in the 0.2 M HCl pool.
3.2 Ecosystem base cation mass-balance budgets
Inputs of Ca, K, and Na via weathering were 1.8-, 1.5-, and 1.3-fold greater, respectively, than inputs via atmospheric deposition, while deposition inputs of Mg were 2-fold greater than weathering inputs. Leaching losses of Ca and Mg were highest in the downslope Regosol from the groundwater discharge area, and leaching losses were less than the combined inputs of weathering and deposition for Ca, K, and Na, but not for Mg (Table 2). Whole-tree harvesting resulted in considerably greater elemental losses than stem-only harvest (2.3–2.6 fold greater). Whole-tree harvesting resulted in net loss of Ca, K, and Mg on all three plots, while stem-only harvesting resulted in net losses of only Mg on all three plots and net losses of Ca on the downslope Regosol (Table 2).
Table 2Mass balances for the three soils collected along the hydrological gradient at Kindla based on annual fluxes of Ca, Mg, K, and Na (mg m−2 yr−1).
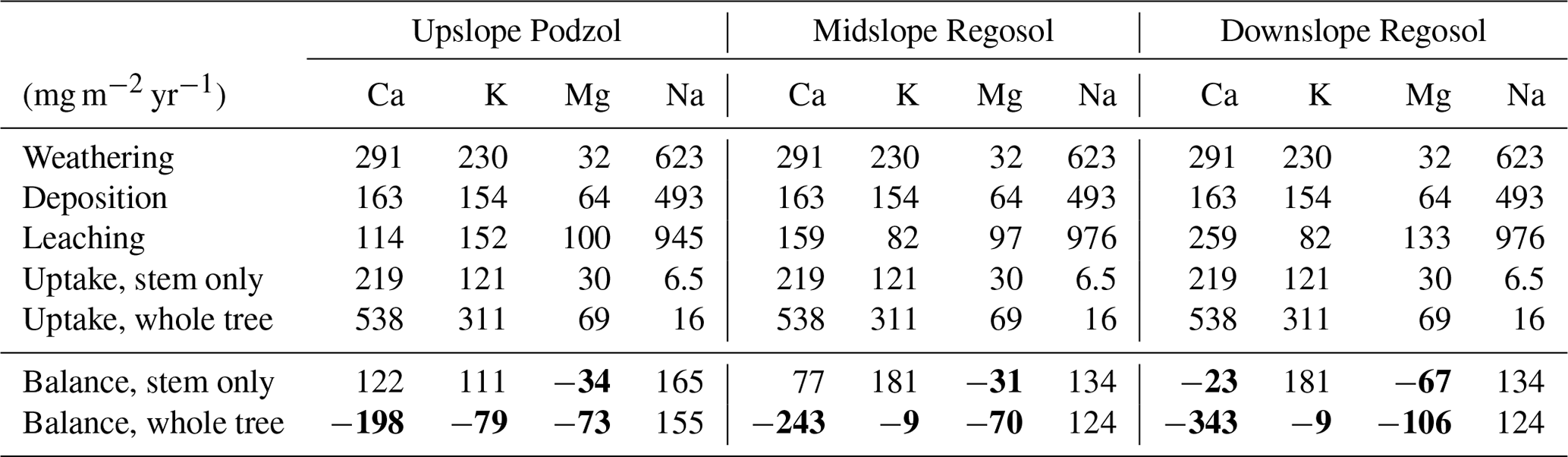
Mass balances in bold are negative, indicating a net loss of base cations from the system.
3.3 Base cation depletion over time
For each harvest scenario–element-soil combination under which a net ecosystem loss was predicted, we calculated the number of 65-year harvest rotations that each extractant-defined base cation pool could offset. Based solely on the mineral soils, the exchangeable (BaCl2-extractable) pools of Mg were not sufficient to sustain more than a single 65-year rotation of stem-only or whole-tree harvest (Table 3). Exchangeable Ca pools were sufficient to sustain 0.3, 0.9, and 1.1 rotations, and K pools were sufficient to sustain 1.2, 4.0, and 6.6 rotations of whole-tree removal on the upslope, midslope, and downslope soils, respectively (Table 3). If the humus layer BaCl2-extractable base cation pools are also considered, the number of rotations of whole-tree harvest which could be sustained by the BaCl2-extractable Mg or Ca pools remains less than two for all hillslope locations. When the humus layer BaCl2-extractable K pools are also considered, the number of rotations which could be sustained by the BaCl2-extractable K pool increases along the hillslope gradient to 2.9, 17, and 33 65-year rotations (Table 3). If they were available for plant uptake, the 0.2 M HCl-extractable base cation pools would be able to sustain considerably more rotations of net ecosystem losses predicted under whole-tree harvesting than the BaCl2-extractable pools. The mineral soil 0.2 M HCl-extractable pools of Mg were sufficient to sustain at least 66 rotations and the 0.2 M HCl-extractable pools of Ca at least 5, rotations of whole-tree removal. The 0.2 M HCl-extractable pools of K pools were sufficient to sustain 19, 137, and 139 rotations of whole-tree removal on the upslope, midslope, and downslope soils, respectively (Table 3). The number of rotations that could be sustained by the 0.5 M HCl-extractable base cation pools was 15 %–90 % greater than by the 0.2 M HCl-extractable pools, while the total amounts of base cations, determined by the aqua regia extraction, could sustain at least 38 whole-tree harvesting rotations (Table 3).
Table 3Number of forest harvest rotations (65 years) each base cation pool may offset net ecosystem losses for each of the three soils. All extractions were performed on the mineral soil. For BaCl2 and aqua regia extractions, humus samples from nearby soils were separately sampled and analyzed, and these humus-derived pools have been added to the mineral soil pools for comparison.
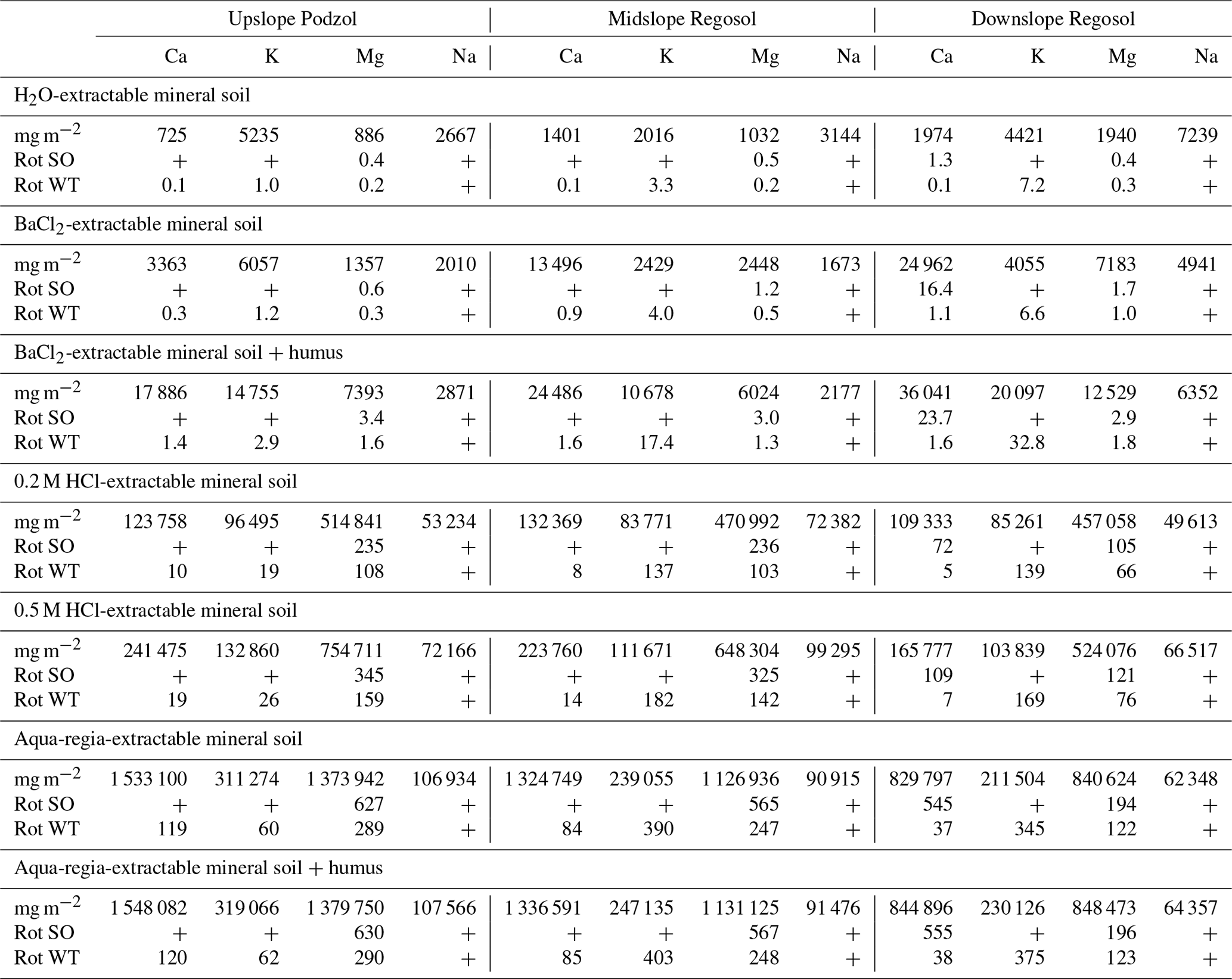
“+” indicates no net loss. “Rot SO” is the number of 65-year rotations of stem-only harvest. “Rot WT” is the number of 65-year rotations of whole-tree harvest.
3.4 Potential sources of acid-extractable base cations in the mineral soil
Given the large size and potential to buffer against leaching and many forest harvest rotations of net base cation loss, understanding the chemical nature and availability of the HCl-extractable pool is important to forming management recommendations. Due to lack of data from the organic horizon, this chapter is primarily focused on the results from the mineral soils. Extractions with strong acids at moderate (0.05 M–1 M) concentrations, as was performed in this study, have been used to selectively extract calcium oxalate (Dauer and Perakis, 2014; Cromack et al., 1979), apatite (Blum et al., 2002; Nezat et al., 2007), non-exchangeable clay interlayer K (Simonsson et al., 2016; Li et al., 2015), and Fe and Mn oxides (Krasnodebska-Ostrega et al., 2001) from soils.
Despite the prevalence of their use, the use of HCl and HNO3 at room temperature or above, from 0.01 to 2 M strength, as extractants, are not specific to particular chemical compounds or soil minerals as all primary and likely all secondary minerals and organocomplexes of Ca, K, Mg, and Na are susceptible to proton-promoted dissolution (Skyllberg et al., 2001), albeit to varying degrees. Because these acid extractions solubilize a range of compounds from mineral soils, elucidating the predominant physicochemical origin of the base cations in the very large HCl-extractable portion is challenging. It would aid our understanding of the potential availability and flux rates of base cations to know, for example, whether the HCl-extractable pool is comprised of incompletely weathered minerals, either as weathering rinds remaining after non-stoichiometric dissolution or secondary mineral coatings that form during weathering, or if it is comprised of secondary compounds that formed after base cations have entered the soil solution via weathering reactions (and may thus also be fed by plant-derived base cations). By examining the base cation ratios and amounts in different extractions we may be able to draw some conclusions about the origins and from which pools the sizable HCl-extractions of base cations derive.
3.4.1 Microbial biomass
Microbial biomass has been suggested to be an important reservoir of N and P, and potentially of base cations. We estimated the potential size of the mineral soil microbial biomass base cation pools as a function of soil carbon and microbial biomass carbon to nutrient ratios (for more detail see Supplement methods). Microbial biomass was estimated to constitute a small but potentially significant fraction of Ca, Mg, and K pools, particularly K. Excluding the humus layer, which is dominated (88 %–100 %) by the BaCl2-extractable base cation pools, the potential microbial biomass pool of Ca was 5 %–36 % as large as the BaCl2-extractable pool, and the potential microbial biomass pool of K was considerably larger than the BaCl2-extractable K pool (by a factor of 1.2–4.2) for the two lower Regosol soils and 26 %–90 % of the BaCl2-extractable K pool at the upper Podzol site. If we assume that microbial biomass contents were extracted in the 0.2 M HCl and stronger acid extractions, microbial biomass constituted on average 2 % and 16 % of 0.2 M HCl-extractable Ca and K pools, respectively. In the downslope Regosol, which lies in a groundwater discharge area and had much higher organic carbon contents, microbial biomass may have contributed 4 % and 26 % of 0.2 M HCl-extractable Ca and K pools, respectively (Fig. 2; Table 4).
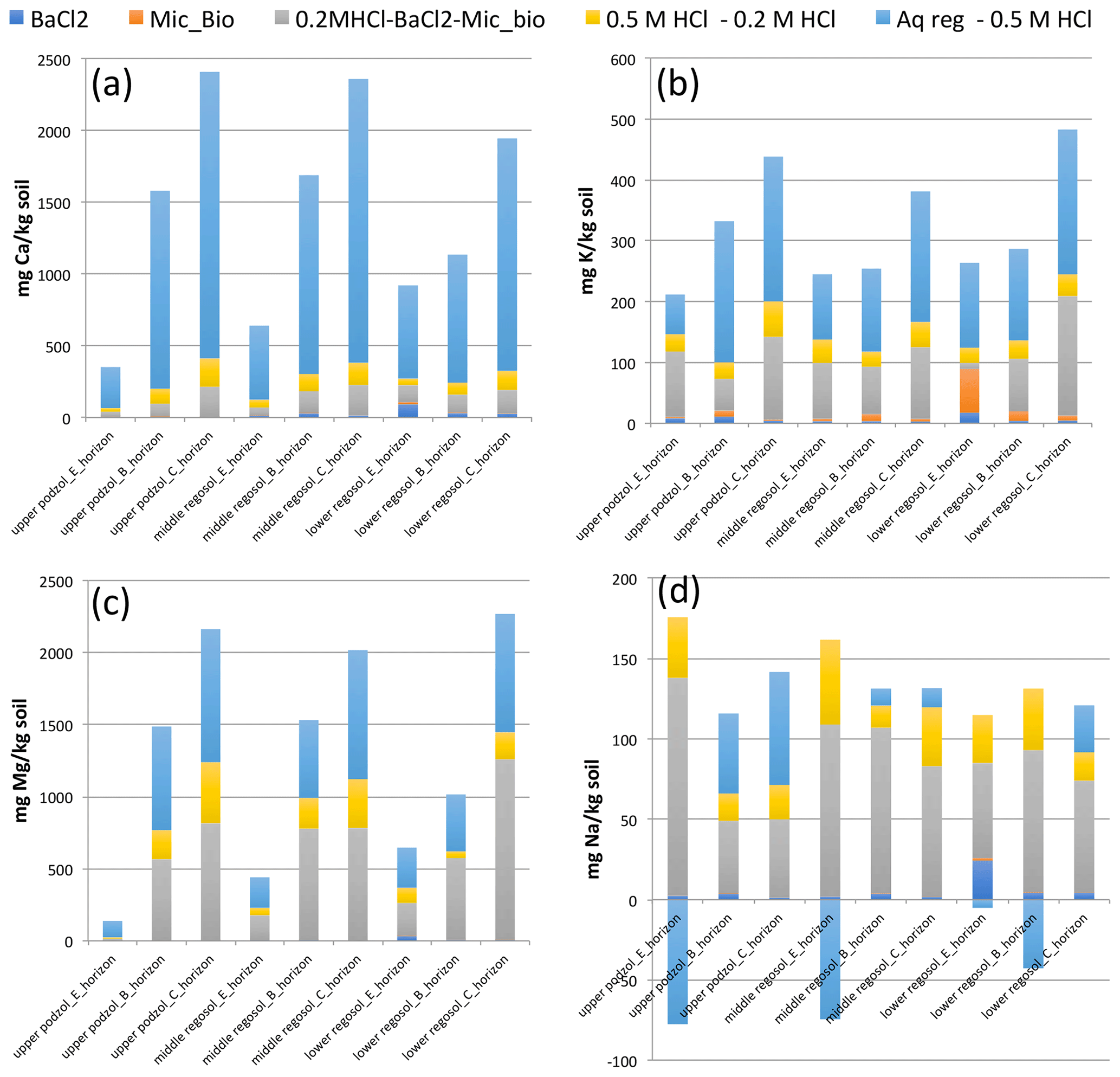
Figure 2Base cation pool sizes of calcium (a), potassium (b), magnesium (c), and sodium (d) across three soils and three soil horizons obtained from soil extraction (BaCl2 extractions), differences between extractions (aqua regia, 0.5 and 0.2 M HCl extractants) and extrapolated estimates (Mic_Bio – microbial biomass; estimated from soil organic carbon contents).
Table 4Mineral soil base cation concentrations from different soil fractions as defined by extractant and relative molar fraction of each base cation within each extraction.

3.4.2 Differences between acid-extracted base cation pools
If we assume that both the 0.2 and 0.5 M HCl extractions extract all, or nearly all, of the BaCl2-extractable and microbial biomass fractions in mineral soil, and if we assume that the aqua regia extraction will also extract all the base cations that the 0.5 M HCl extraction would, then we can examine the differences between these extractions as being more diagnostic of the mode of chemical attack of the more aggressive (in terms of greater extraction yield) extractant. We will use the term “only” to designate base cation pool estimates that are derived from subtracting extracted amounts from one extractant pool from another. Thus, the 0.2 M HCl-only pool is what remains of the 0.2 M HCl-extractable base cation pools after both the estimated microbial biomass and BaCl2-extractable base cation pools were subtracted; 0.5 M HCl-only is the difference between the 0.5 and 0.2 M HCl-extractable pools; aqua regia only is the difference between the aqua-regia- and 0.5 M HCl-extractable pools. We can further examine the molar ratios of base cations in these pools and compare both BC pool sizes and elemental stoichiometry as well as examine potential relationships between these and clay and organic matter contents (Table 1).
The pool of aqua-regia-only Ca is far larger (4–5 fold) than either of the HCl-only-extractable Ca pools (Table 4; Fig. 2). This stands in contrast to K, Mg, and Na, for which the 0.2 M HCl-only pools are nearly as large or larger than the aqua-regia-only fraction. Calcium also stands out in that the 0.5 and 0.2 M HCl-only digestible fractions were similar in size while, in contrast, for K, Mg, and Na the stronger acid yielded relatively less additional cations (less than half as much as the 0.2 M HCl only). Calcium is the dominant cation (comprising 50 % or more of total base cations, on a molar basis) in both the exchangeable and aqua regia base cation pools, but in the HCl-extractable pools it comprised a smaller portion (Table 4). In contrast we see for Mg that the two HCl-only pools combined are larger than aqua-regia-only pools of Mg, and we also see that the mole fraction of base cations is heavily enriched for Mg in the HCl-extractable fraction. HCl-extractable Mg also displays a strong depth trend in each of the three soils, being 5–70 times as abundant in the C-horizon HCl-only extracts as compared to E-horizon extracts. While the amounts of both Mg and Ca increase with depth in the HCl extractions, HCl-extractable K does not display the same behavior; in the upslope and midslope soils its abundance is relatively stable with depth, and the relative abundance of K ([K]∕[BCtot]) is notably higher in the uppermost E horizon.
3.4.3 Base cations bound to soil organic matter
Base cations may be bound to organic compounds in the soil and organically bound base cations may represent a significant pool of exchangeable cations (Duchesne and Houle, 2008; Richardson et al., 2017). The humus layer 0.1 M BaCl2-extractable pools of Ca, K, Mg, and Na were 88 %, 100 %, 86 %, and 99 %, respectively, as large as the humus-layer aqua-regia-extracted pools. Focusing on the divalent ions calcium and magnesium, these results as well as earlier studies (Bailey et al., 2003; Dauer and Perakis, 2013) show that a significant fraction of the organically bound cations may not be extractable with concentrated salts. Calcium, for example, may form strong complexes with organic acids (Tipping, 2002), such as calcium oxalate. Calcium oxalate has been suggested to accumulate in soils over time, potentially in large amounts (Cromack et al., 1979; Dauer and Perakis, 2013). If organically bound calcium were a significant portion of the HCl-extractable pools, we might expect to see a decrease with depth as Ca oxalate is mostly associated with exudation and incomplete decomposition of organic matter near the soil surface. In addition we might expect to see a relative increase in the Ca:BCtot elemental ratios, especially near the soil surface. Instead we observed that the Ca:BCtot ratios are relatively depleted in the HCl-extractable fractions compared to both the exchangeable and aqua-regia-extractable fractions; in addition, we see no change or a slight increase in the Ca:BCtot ratios with depth in the HCl-extractable pools. Moreover, we would expect organically bound Ca to be largely dissolved by the 0.2 M HCl extraction, and thus if it dominated the HCl-only pools, we would expect considerably less Ca in the 0.5 M HCl-only pool, which we do not find. Finally, if a significant pool of the acid-extractable base cations were locked in organic matter, we would expect a correlation between soil carbon and the acid-extractable BC pools, but we do not. We do see a fairly strong correlation between organic carbon and BaCl2-extractable Ca, Mg, and Na pools (%C vs. Ca, r2=0.56; vs. K, r2=0.00; vs. Mg, r2=0.61; vs. Na, r20.71; data not shown), but not between organic C and acid-extractable BC pools. Taken together these observations suggest that organically bound Ca (including Ca oxalate) is unlikely to be a major component of the HCl-extractable Ca pools. Indeed, Dauer and Perakis (2014) observed that while production rates of Ca oxalate may be high, Ca oxalate is also highly susceptible to microbial degradations, such that acid-extractable Ca oxalate pools comprised less than 3 % of exchangeable Ca pools.
Soil organic matter (SOM) may also form protective coatings on soil minerals reducing their solubility (Drever and Stillings, 1997). It is possible that a significant portion of mineral surfaces were shielded with SOM, and the HCl extractions may have removed these protective layers. This could potentially explain why, particularly for K and Mg, we see considerably greater BC yields in the 0.2 M HCl-only fraction than we see in the 0.5 M HCl-only fraction because 0.2 M HCl is sufficient to remove these protective coatings, and the resultant burst of mineral surface dissolution is not a function of acid strength but a function of the removal of these coatings. To examine the potential for organic matter coatings on minerals to shield them from weathering but be readily removed by 0.2 M HCl, chemical treatments utilizing hydrogen peroxide or surfactants (Chao, 1984) could be used to remove or reduce such coatings without the use of a strong acid.
3.4.4 Clay interlayer potassium
Non-structural K, strongly bound in clay interlayers has been observed to be present in large quantities in a variety of soils (Moritsuka et al., 2003; Li et al., 2015). Clay interlayer K is typically extracted with dilute or concentrated HNO3 or HCl, or specific cation-exchange reactants, such as sodium tetraphenylboron (NaTPB). HCl and HNO3 have been shown to release significant portions of this pool, but also to cause significant, though limited, weathering of clays; NaTPB, in contrast has been shown to release far more interlayer K and not exhibit any observable weathering (Li et al., 2015; Moritsuka et al., 2003). If clay interlayer exchangeable or occluded K were a major storage pool, then we might expect the size of the acid-extractable K pool to correlate with clay content, which we do not see. Instead, there is a weak negative correlation between both the 0.2 and 0.5 M HCl-extractable K and clay content (as measured by particle size, Table 1). However clay content, as measured by particle size may not accurately reflect the amount of secondary clay minerals, or, in particular, the amount of minerals which are likely reservoirs of interlayer K (Simard et al., 1989). Indeed, Mengel and Rahmatullah (1994) observed that the clay interlayer K pool was both large and increasingly important for plant uptake in coarser soil mixtures as clay content decreased, due to the relative reduction in exchangeable K, and Simonsson et al. (2016) showed that clay interlayer K may constitute a very large pool of K (> 20 × exchangeable K) even when clay contents were quite low (3 %–5 %). We observed that HCl-extractable K in the E horizon was larger than that of the B horizon and similar in size to the C horizon in two of the three soils (upslope and midslope). This lack of a clear depth trend, which both Mg and Ca have, in conjunction with the abovementioned high proportion of the total base cation pool which is K in the E horizon, may indicate an accumulation of non-structural clay interlayer K in the upper horizons of the upslope and midslope soils. Use of NaTPB, instead of HCl, or use of repeated sequential extractions of cold 0.01 M HCl (Moritsuka et al., 2003) may avoid the non-specific dissolution activity of stronger acids and be more diagnostic for extraction of K bound in clay interlayers.
3.4.5 Fe and Mn oxides
HCl is commonly used, both as a single extractant and in sequential extractions to dissolve Fe and Mn oxides in soils (Chao and Sanzolone, 1992; Krasnodebska-Ostrega et al., 2001). Soil Fe and Mn oxides can be important reservoirs for base cations not extractable with typical salt-exchange assays (Krasnodebska-Ostrega et al., 2001). Indeed we extracted large amounts of Fe in the 0.2 M HCl-only pool (> 50 % total Fe, 0.01 %–0.1 % total soil dry mass), though relatively little Mn (th as much as Fe by mass; data not shown). There was no significant correlation between Fe extracted in any of the acid fractions and the contents of any base cations in those fractions. In addition, the HCl-only pools had the lowest BCtot:Fe ratios amongst all extraction pools (data not shown). Our data indicate that despite being both readily extractable and present in the soils in very high amounts, binding to Fe oxide fractions appears unlikely to account for the majority of the acid-extractable base cation pools.
3.4.6 Primary minerals
While more aggressive digestions (aqua regia, HF, LiBO2 fusion) are typically used to estimate elemental contents of primary minerals in soils, the large pools of BC observed in the HCl digestions may have largely arisen from the HCl-induced dissolution of primary minerals. The Ca, K, and Mg contents of the 0.2 M HCl-only pools correspond to 357, 174, and 6832 years of mineral weathering, given the weathering rates used in this study from McGivney et al. (2019). If this dissolution were congruent and in proportion to the soil mineral contents, we might expect similar elemental ratios in the HCl-only-extractable pools as we observe in the aqua regia pool. We see instead that Ca:BCtot are relatively lower, and K:BCtot ratios and Mg:BCtot ratios are relatively higher in the HCl-extractable fraction as compared to the aqua regia digestion. However, incomplete mineral dissolution with HCl and HNO3 at concentrations of 1 M or less is known to yield incongruent dissolution, preferentially releasing base cations from primary minerals (Snäll and Liljefors, 2000; Chao, 1984; Moritsuka et al., 2003) and sediments (Agemian and Chau, 1976; Sutherland, 2002) and the distribution of minerals in different soil particle size classes is understood not to be representative of bulk mineralogy (Simard et al., 1989). The size, depth distribution and relatively higher molar ratio of Mg in the HCl-only pool support the possibility of easily weatherable Mg-containing minerals as a likely source. Modeled mineralogy from total soil elemental contents in the Kindla soils indicate the presence of apatite (0.1 %–0.2 %), chlorite (0.2 %–0.8 %), and hornblende (0.3 %–1.8 %) in the E and B horizons (McGivney et al., 2019), and regional databases indicate the presence of these minerals as well as biotite in soils with similar geologic origin (Stendahl et al., 2002). In mineral and soil dissolution studies chlorite, hornblende, and biotite have been shown to yield high quantities of Mg in extractions with 1 M HCl (Snäll and Liljefors, 2000), although Mg extraction efficiency from hornblende were considerably lower and more particle size dependant. For Ca, apatite stands out as a likely mineral source of the HCl-extractable Ca, but were apatite the dominant source of Ca, we might expect a different relative distribution between the different extractions. As apatite is highly susceptible to proton-promoted dissolution, more so than the aforementioned potential Mg source pools (Nezat et al., 2007), we would expect relatively larger portions of total Ca to have been extracted in the 0.2 M HCl-only vs. 0.5 M HCl-only pools and in the HCl-only vs. the aqua regia pools than we see. These findings indicate that either significant portions of the Ca are coming from more recalcitrant minerals than apatite, or that there is significant mineral shielding of apatite, and stronger acids than 0.2 M HCl are needed to scavenge these apatite inclusions. Nezat et al. (2007) observed that the 70 % of apatite mineral originally present in the soil was weathered completely in the upper soil layers and the remaining 30 % was shielded by inclusion in granitoid minerals and thus inaccessible to acid digestion with 2 M HNO3 (though accessible to acid digestion with HF). Were apatite to be a primary source of Ca in the 0.2 and 0.5 M HCl-only extractions we might expect Ca:P molar rations close to 10:6 as is the case for apatite. Using this ratio, assuming congruent dissolution, and assuming all of the P contents in the 0.5 M HCl-only extraction could be attributable to apatite, 20 %–27 % of the C-horizon Ca content in 0.5 M HCl-only extractions could be attributable to apatite.
3.4.7 Physicochemical origin of HCl-extractable base cation pools
Our data indicate that the large pools of 0.2 M HCl-extractable base cation pools may be comprised of a combination of microbial biomass, the dissolution of fine grain, potentially OM-shielded, easily weatherable minerals (such as biotite, apatite, chlorite, and hornblende), as well as by the presence of significant stores of non-structural clay interlayer bound K. Determining the physicochemical nature of these base cation pools is central to incorporating them into mass-balance and geochemical models widely used today to inform management and policy. If these base cation pools derive from primary mineral weathering, then weathering models may already account for these pools or may need to be adjusted to account for these large and potentially labile pools; if they are a result of organic complexation or secondary minerals, then understanding what controls their formation and availability depends on a better understanding of their chemical nature. Combining sequential extraction approaches with X-ray diffraction methods on small soil samples of different particle size classes may help elucidate the potential importance of primary minerals to the HCl-extractable pools, while other spectroscopic methods (e.g., micro, nanoSIMS, STXM) may be employed to determine the role of organic or secondary mineral coatings or identify secondary mineral or non-crystalline base cation pools.
Among the many studies which have predicted net ecosystem losses of base cations with biomass (stem only or whole tree) extractions, exchangeable base cation pools, when measured, are commonly not sufficient to sustain more than a single harvest rotation, assuming productivity, growth, and base cation uptake are not reduced by reductions in base cation availability (Akselsson et al., 2007a, b; Duchesne and Houle, 2008; Knust et al., 2016). We are not aware of any other studies of ecosystem mass balance of base cations that have attempted to explore the sizes of non-exchangeable nutrient pools in relation to net ecosystem losses over time. Those that have compared HCl- or HNO3-extractable base cation contents in similar soils have commonly found acid-extractable pools to be many fold larger than salt-extractable exchangeable pools (Olofsson et al., 2016; Nezat et al., 2007; Simard et al., 1989; Moritsuka et al., 2002; Lucash et al., 2012).
The relevance of the size of these large pools of acid-extractable base cations to forest nutrient relations is a function of their flux rates or bioavailability. If the base cations released from 0.2 M HCl-only pools are highly stable over centuries and not available to refill depleted exchange sites, from which plant uptake may occur, then they are of little relevance to policy decisions on sustainable harvest levels. If, on the other hand, they are in equilibrium with the exchangeable pools and available for plant uptake in significant amounts on decadal or century timescales then these pools and their size relative to the exchangeable pool are of immediate relevance to forest management recommendations in light of the immediate need to reduce fossil fuel emissions.
A number of studies have examined the bioavailability and flux into exchangeable pools of acid-extractable non-exchangeable pools of base cations across a variety of soils. Callesen et al. (2004) examined 19 Danish forest soils comparing exchangeable base cation amounts to base cations pools which were extractable with 0.01 M HNO3 and conducted repeated sequential HNO3 extractions over periods of hours to days. They observed that base cation pools extracted with 0.01 M HNO3 in 2 h were similar in magnitude to BaCl2-exchangeable pools but that in repeated extractions with HNO3, similar amounts of base cations continued to be extractable, while the size of BaCl2-exchangeable pools decreased markedly after the first extraction. Morisuka et al. (2002) observed discreet patterns of bioavailability of the dilute 0.01 M HCl-extractable potassium pool from the exchangeable pool; during the course of a 17 d growth experiment with Zea mays, both soil pools contributed to plant K, but the HCl-extractable pool exhibited much shorter depletion zones around mays roots than the exchangeable pool, suggesting a more active role by plants in uptake from the acid-extractable pool. Ortas et al. (1999) grew Italian grass in pots for 6 months and measured the exchangeable and acid-extractable K and Mg contents of the soil before and after; they observed that, on average, ∼45 % and ∼20 % of plant K and Mg contents, respectively, were derived from the acid-extractable, non-exchangeable soil pools. Mengle and Rahmatullah (1994) grew a number of crops in soils of varying exchangeable K availability and observed that on the soils poorest in exchangeable K, the majority of plant K derived from acid-extractable non-exchangeable pools. Taken together these and other findings clearly indicate that base cation pools that are extractable with moderately concentrated strong acids, and not extractable with salt solutions used to assay the exchangeable base cation pools, may be readily available for plant uptake and in some equilibrium with exchangeable pools. However, more research is needed to determine what controls the availability of acid-extractable non-exchangeable pools and the rate of flux into exchangeable pools.
Simonsson et al. (2016) used K:Rb ratios in different soil extracts to examine the flux rates between HCl-extractable putatively clay interlayer K and exchangeable K and concluded that there was active and continuous exchange between the exchangeable and the “non-exchangeable” HCl-extractable K pools during a single forest rotation. A number of studies have used stable or radio-isotope tracers in plant–soil systems to examine the relative contributions of different soil pools to plant uptake. Blum et al. (2002) used Ca:Sr and 87Sr:86Sr isotope ratios to conclude that apatite minerals were a particularly important source of Ca to ectomycorrhizal trees colonized by ectomycorrhizal fungi. Isotopic dilution experiments provide a promising tool to quantify source and sink reservoirs in the soil and estimate the rates of elemental flux between different soil pools. Newbould and Russell (1963) observed that 45Ca added to pots equilibrated rapidly with the exchangeable Ca in the soil, but when they grew ryegrass in those pots, plants accessed additional Ca from a pool that was not exchangeable. The isotopic dilution of 44Ca and 26Mg tracers added to a Ca- and Mg-poor beech forest in central France showed that there may be important Ca sources in the soil that are not accounted for by conventional mass-balance approaches (van der Heijden et al., 2014). Isotopic dilution experiments carried out in the lab on unlabeled soil from the same beech forest showed that the oxalic acid-extractable and the HNO3-extractable pools contributed directly to the labile pools of Mg, Ca, and K. They concluded that these acid-extractable, non-exchangeable pools of Mg, Ca, and K are likely to be significant to nutrient cycling on the scale of years to decades. Though isotopic approaches enable the study of elemental fluxes between extractable soil pools, but they are insufficient to better characterize the physicochemical nature of these pools. As discussed above, this challenging question requires the development and combination of new methods such as spectroscopic methods (e.g., micro, nanoSIMS, STXM) to better understand what organic, mineral, and organomineral phases are targeted by the different extraction reactants in order to determine the chemical nature of these pools.
Our mass-balance estimates indicate that stem-only harvesting would moderately deplete and whole-tree harvesting would markedly deplete exchangeable cation pools, such that current 0.1 M BaCl2-exchangeable pools of base cations are insufficient to support more than a single forest rotation under whole-tree harvesting of the Kindla forest. A large fraction of these salt-extractable base cation pools were found in the organic rich horizons, especially the humus layer. HCl-extractable base cation pools are much larger than the salt-exchangeable pools, and if available for plant uptake and/or exchange with the exchangeable pool they could sustain many rotations of stem-only or whole-tree harvesting. The large pools of 0.2 M HCl-extractable base cations we observed may be comprised of a combination of microbial biomass, fine-grained easily weatherable minerals that may be protected from weathering by organic matter or secondary mineral coatings, as well as by the presence of significant stores of non-structural clay interlayer bound K. We were, however, unable to apportion the HCl-extractable base cation pool between these potential pools or draw any conclusions about their flux rates or bioavailability. Specific extraction techniques, including H2O2 or other oxidative treatments that are specific to organic matter, sodium tetraphenylboron to extract clay interlayer K without dissolving minerals, as well as more dilute (0.01 M) acid extractions, may help us to more diagnostically estimate the size of different base cation pools. Spectroscopic techniques may allow us to identify the chemical nature of these pools, while isotopic techniques may allow the quantification of base cation flux rates between exchangeable and acid-extractable pools.
Many soils appear to have very large, relative to exchangeable pools, base cation reserves in the mineral soil, which are not extractable with conventional exchangeable cation assays but which appear to be available for plant uptake and interact with the exchangeable pool on the scale of years to decades. These large pools should be addressed in ecosystem mass-balance research and accounted for and considered in nutrient management recommendations. To develop forest management policy based on these putatively available pools, a better understanding of their physicochemical nature, bioavailability and flux rates is needed.
Data are stored at the Department of Aquatic Sciences and Assessment at the Swedish University of Agricultural Sciences and are made available on request by stefan.lofgren@slu.se or lars.lundin@slu.se.
The supplement related to this article is available online at: https://doi.org/10.5194/soil-5-351-2019-supplement.
NPR, KB, and SL conceived the idea for the paper. LL, SL, and EM contributed data. NPR analyzed the data and wrote the paper. All authors contributed ideas, participated in discussions about the paper, and were involved with writing and interpretation.
The authors declare that they have no conflict of interest.
This article is part of the special issue “Quantifying weathering rates for sustainable forestry (BG/SOIL inter-journal SI)”. It is not associated with a conference.
The authors would like to thank the following funding agencies responsible for this work. The Swedish Research council for funding the research consortium: Quantifying weathering Rates for Sustainable Forestry; the Swedish Energy Agency; the European Commission; and the International Cooperative Programme on Assessment and Monitoring of Air Pollution Effects on Forests (ICP Forests) for providing support for data collection and for the SoilTrEC project.
This research has been supported by the Svenska Forskningsrådet Formas (grant no. 212-2011-1691).
This paper was edited by Boris Jansen and reviewed by two anonymous referees.
Achat, D. L., Deleuze, C., Landmann, G., Pousse, N., Ranger, J., and Augusto, L.: Quantifying consequences of removing harvesting residues on forest soils and tree-growth – a meta-analysis, Forest Ecol. Manag., 348, 124–141, 2015.
Agemian, H. and Chau, A. S. Y.: Evaluation of Extraction Techniques for the Determination of Metals in Aquatic Sediments, The Analyst, 101, 761–767, 1976.
Aherne, J., Posch, M., Forsius, M., Lehtonen, A., and Härkönen, K.: Impacts of forest biomass removal on soil nutrient status under climate change: a catchment-based modelling study for Finland, Biogeochemistry, 107, 471–488, https://doi.org/10.1007/s10533-010-9569-4, 2012.
Akselsson, C. and Belyazid, S.: Critical biomass harvesting – Applying a new concept for Swedish forest soils, Forest Ecol. Manag., 409, 67–73, https://doi.org/10.1016/j.foreco.2017.11.020, 2018.
Akselsson, C., Westling, O., Sverdrup, H., Holmqvist, J., Thelin, G., Uggla, E., and Malm, G.: Impact of harvest intensity on long-term base cation budgets in Swedish forest soils, Water Air Soil Pollut. Focus, 7, 201–210, https://doi.org/10.1007/s11267-006-9106-6, 2007a.
Akselsson, C., Westling, O., Sverdrup, H., and Gundersen, P.: Nutrient and carbon budgets in forest soils as decision support in sustainable forest management, Forest Ecol. Manag., 238, 167–174, https://doi.org/10.1016/j.foreco.2006.10.015, 2007b.
Bailey, S. W., Buso, D. C., and Likens, G. E.: Implications of sodium mass balance for interpreting the calcium cycle of a forested ecosystem, Ecology, 84, 471–484, 2003.
Banwart, S., Bernasconi, S., Bloem, J., Blum, W., Brandao, M., Brantley, S., Chabaux, F., Duffy, C., Lundin, L., Kram, P., Nikolaidis, N., Novak, M., Panagos, P., Ragnarsdottir, K.V., Reynolds, B., Rousseva, S., de Ruiter, P., van Gaans, P., van Riemsdijk, W., White, T., and Zhang, B.: Soil processes and functions in critical zone observatories: Hypothesis and experimental design. Special section: Critical Zone Observatories, Vadose Zone J., 10, 974–987, 2011.
Blum, J. D., Klaue, A., Nezat, C. A., Driscoll, C. T., Johnson, C. E., Siccama, T. G., Eagar, C., Fahey, T. J., and Likens, G. E.: Mycorrhizal weathering of apatite as an important calcium source in base-poor forest ecosystems, Nature, 417, 729–731, 2002.
Brandtberg, P. O. and Olsson, B. A.: Changes in the effects of whole-tree harvesting on soil chemistry during 10 years of stand development, Forest Ecol. Manag., 277, 150–162, 2012.
Callesen, I. and Raulund-Rasmussen, K.: Base cation, aluminum, and phosphorus release potential in Danish forest soil, J. Plant Nutr. Soil Sc., 167, 169–176, 2004.
Chao, T. T.: Use of partial dissolution techniquesin geochemical exploration, J. Geochem. Explor., 20, 101—135, 1984.
Chao, T. T. and Sanzolone, R. F.: Decomposition techniques, J. Geochem. Explor., 44, 65–106, 1992.
Cools, N. and de Vos, B.: Sampling and Analysis of Soil, Manual Part X, in: Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests, UNECE, ICP Forests, Hamburg, ISBN: 978-3-926301-03-1, http://www.icp-forests.org/Manual.html (last access: 4 April 2019), 208 pp., 2010.
Cromack, K., Sollins, P., Graustein, W. C., Speidel, K., Todd, A. W., Spycher, G., Li, C. Y., and Todd, R. L.: Calcium oxalate accumulation and soil weathering in mats of the hypogeous fungus Hysterangium crassum, Soil Biol. Biochem., 11, 463–468, 1979.
Dauer, J. M. and Perakis, S. S.: Contribution of Calcium Oxalate to Soil-Exchangeable Calcium, Soil Sci., 178, 671–678, https://doi.org/10.1097/SS.0000000000000029, 2013.
Dauer, J. M. and Perakis, S. S.: Calcium oxalate contribution to calcium cycling in forests of contrasting nutrient status, Forest Ecol. Manag., 334, 64–73, 2014.
de Oliveira Garcia, W., Amann, T., and Hartmann, J.: Increasing biomass demand enlarges negative forest nutrient budget areas in wood export regions, Sci. Rep., 8, 5280, https://doi.org/10.1038/s41598-018-22728-5, 2018.
Drever, J. I. and Stillings, L. L.: The role of organic acids in mineral weathering, Colloids Surf A, 120, 167–181, 1997.
Duchesne, L. and Houle, D.: Impact of Nutrient Removal through Harvesting on the Sustainability of the Boreal Forest, Ecol. Appl., 18, 1642–1651, 2008.
Duval, B. D., Blankinship Djikstra, P., and Hungate, B. A.: CO2 effects on plant nutrient concentration depend on plant functional group and available nitrogen: A meta-analysis, Plant Ecol., 213, 505–521, 2012.
Falk Øgaard, A. and Krogstad, T.: Release of interlayer potassium in Norwegian grassland soils, J. Plant Nutr. Soil Sc., 168, 80–88, 2005.
Feller, M. C.: Forest harvesting and streamwater inorganic chemistry in Western North America: A review, J. Am. Water Resour. As., 41, 785–811, 2005.
Grove J. H., Sumner M. E., and Syers J. K.: Effect of Lime on Exchangeable Magnesium in Variable Surface Charge Soils, Soil Sci. Soc. Am. J., 45, 497–500, 1981.
Hall, S. J. and Huang, W.: Iron reduction: a mechanism for dynamic cycling of occluded cations in tropical forest soils?, Biogeochemistry, 136, 91–102, 2017.
Hamburg, S. P., Yanai, R. D., Arthur, M. A., Blum, J. D., and Siccama, T. G.: Biotic control of calcium cycling in northern hardwood forests: acid rain and aging forests, Ecosystems, 6, 399–406, 2003.
Iwald, J., Löfgren, S., Stendahl, J., and Karltun, E.: Acidifying effect of removal of tree stumps and logging residues as compared to atmospheric deposition, Forest Ecol. Manag., 290, 49–58, 2013.
Kazagic, A., Music, M., Smajevic, I., Ademovic, A., and Redzic, E.: Possibilities and sustainability of “biomass for power” solutions in the case of a coal-based power utility, Clean Technol. Envir., 18, 1675–1683, 2016.
Kinniburgh, D. G., Jackson, M. L., and Syers, J. K.: Adsorption of Alkaline Earth, Transition, and Heavy Metal Cations by Hydrous Oxide Gels of Iron and Aluminum, Soil Sci. Soc. Am. J., 40, 796–799, 1976.
Knust, C., Schua, K., and Feger, K. H.: Estimation of Nutrient Exports Resulting from Thinning and Intensive Biomass Extraction in Medium-Aged Spruce and Pine Stands in Saxony, Northeast Germany, Forests, 7, 302, https://doi.org/10.3390/f7120302, 2016.
Krasnodebska-Ostrega, B., Emons, H., and Golimowski, J.: Selective leaching of elements associated with Mn – Fe oxides in forest soil, and comparison of two sequential extraction methods, Fresenius J. Anal. Chem., 371, 385–390, https://doi.org/10.1007/s002160100982, 2001.
Kreutzweiser, D. P., Hazlett, P. W., and Gunn, J. M.: Logging impacts on the biogeochemistry of boreal forest soils and nutrient export to aquatic systems: A review, Environ. Rev., 16, 157–179, 2008.
Lauri, P., Havlík, P., Kindermann, G., Forsell, N., Böttcher, H., and Obersteiner, M.: Woody biomass energy potential in 2050, Energ. Policy, 66, 19–31, https://doi.org/10.1016/j.enpol.2013.11.033, 2014.
Li, T., Wang, H., Zhou, Z., Chen, X., and Zhou, J.: A nano-scale study of the mechanisms of non-exchangeable potassium release from micas, Appl. Clay Sci., 118, 131–137, 2015.
Löfgren, S., Aastrup, M., Bringmark, L., Hultberg, H., Lewin-Pihlblad, L., Lundin, L., Karlsson, G., and Thunholm, B.: Recovery of Soil Water, Groundwater, and Streamwater From Acidification at the Swedish Integrated Monitoring Catchments, AMBIO, 40, 836–856, 2011.
Löfgren, S., Ågren, A., Gustafsson, J. P., Olsson, B. A., and Zetterberg, T.: Impact of whole-tree harvest on soil and stream water acidity in southern Sweden based on HD-MINTEQ simulations and pH-sensitivity, Forest Ecol. Manag., 383, 49–60, 2017.
Lorenz, N., Verdell, K., Ramsier, C., and Dick, R. P.: A Rapid Assay to Estimate Soil Microbial Biomass Potassium in Agricultural Soils, Soil Sci. Soc. Am. J., 74, 512–516, 2010.
Lucas, R. W., Holmström, H., and Lämås, T.: Intensive forest harvesting and pools of base cations in forest ecosystems: A modeling study using the Heureka decision support system, Forest Ecol. Manag., 325, 26–36, 2014.
Lucash, M. S., Yanai, R. D., Blum, J. D., and Park, B. B.: Foliar Nutrient Concentrations Related to Soil Sources across a Range of Sites in the Northeastern United States, Soil Sci. Soc. Am. J., 76, 674–683, https://doi.org/10.2136/sssaj2011.0160, 2012.
McGivney, E., Gustafsson, J. P., Belyazid, S., Zetterberg, T., and Löfgren, S.: Assessing the impact of acid rain and forest harvest intensity with the HD-MINTEQ model – soil chemistry of three Swedish conifer sites from 1880 to 2080, SOIL, 5, 63–77, https://doi.org/10.5194/soil-5-63-2019, 2019.
McLaughlin, J. W. and Philips, S. A.: Soil carbon, nitrogen, and base cation cycling 17 years after whole-tree harvesting in a low-elevation red spruce (Picea rubens)-balsam fir (Abies balsamea) forested watershed in central Maine, USA, Forest Ecol. Manag., 222, 234–253, 2006.
Mengel, K. and Rahmatullah: Exploitation of potassium by various crop species from primary minerals in soils rich in micas, Biol. Fertil. Soils, 17, 75–79, 1994.
Moritsuka, N., Yanai, J., and Kosaki, T.: A sensitive method for detecting the depletion of nonexchangealble potassium in the rhizosphere using a sequential extraction with 0.01 molar hydrochloric acid, Soil Sci. Plant Nutr., 48, 577–583, https://doi.org/10.1080/00380768.2002.10409241, 2002.
Moritsuka, N., Yanai, J., Fujii, A., Sano, S., and Kosaki, T.: Evaluation of readily available nonexchangeable potassium in soil by sequential extractions with 0.01 molar hydrochloric acid, Soil Sci. Plant Nutr., 49, 631–639, https://doi.org/10.1080/00380768.2003.10410053, 2003.
Newbould, P. and Russell, R. S.: Isotopic equilibration of calcium-45 with labile soil calcium, Plant Soil, 18, 239–257, 1963.
Nezat, C. A., Blum, J. D., Yanai, R. D., and Hamburg, S. P.: A sequential extraction to determine the distribution of apatite in granitoid soil mineral pools with application to weathering at the Hubbard Brook Experimental Forest, NH, USA, Appl. Geochem., 22, 2406–2421, 2007.
Nilsson, D., Nilsson, B., Thörnqvist, T., and Bergh, J.: Amount of nutrients extracted and left behind at a clear-felled area using the fresh-stacked and dried-stacked methods of logging residue extraction, Scand. J. Forest Res., 33, 437–445, https://doi.org/10.1080/02827581.2018.1427786, 2018.
Nilsson, S. I., Miller, H. G., and Miller, J. D.: Forest Growth as a Possible Cause of Soil and Water Acidification: An Examination of the Concepts, Oikos, 39, 40–49, 1982.
Nilsson, T. and Lundin, L.: Predictions of bulk density of Swedish forest soils from the organic content and soil depth, Report in: Forest Ecology and Forest Soils no 91, Swedish University of Agricultural Sciences, Uppsala, ISSN 0348-3398, 41 pp., 2006 (in Swedish, English summary).
Olofsson, J.: Base cations in forest soils: A pilot project to evaluate different extraction methods, Master thesis, UPTEC W, ISSN 1401-5765, 16031, 51 pp., http://uu.diva-portal.org/smash/record.jsf?pid=diva2%3A1049873&dswid=-3006 (last access: 9 December 2019), 2016.
Olsson, M. and Lundin, L.: Determination of weathering rates and base cation budgets – an example from glacial till in central Sweden, in: Changes to forest management and silvicultural techniques necessitated by forest energy production; Ed: Tham, Å. & Mård, H. Int. Energy Agency/Bioenergy agreement, Activity A1.Conventional Systems, Garpenberg 30 September–4 October 1990, Department of Forest Yield Research, SLU, Report No. 27, 111–122, 1990.
Ortas, I., Güzel, N., and Ibrikçi, H.: Determination of Potassium and Magnesium Status of Soils Using Different Soil Extraction Procedures in the Upper Part of Mesopotamia (in the Harran Plain), Commun. Soil Sci. Plant Anal., 30, 2607–2625, 1999.
Posch, M. and Kurz, D.: A2M – A program to compute all possible mineral modes from geochemical analyses, Comput. Geosci., 33, 563–572, https://doi.org/10.1016/j.cageo.2006.08.007, 2007.
Richardson, J. B., Petrenko, C. L., and Friedland, A. J.: Base cations and micronutrients in forest soils along three clear-cut chronosequences in the northeastern United States, Nutr. Cycl. Agroecosys., 109, 161–179, https://doi.org/10.1007/s10705-017-9876-4, 2017.
Royo, J., Sebastián, F., García-Galindo, D., Gómez, M., and Díaz, M.: Large-scale analysis of GHG (greenhouse gas) reduction by means of biomass co-firing at country-scale: Application to the Spanish case, Energy, 48, 255–267, 2012.
Simard, R. R., de Kimpe, C. R., and Zizka, J.: The kinetics of non-exchangeable potassium and magnesium release from Quebec Soils, Can. J. Soil Sci., 69, 663–675, 1989.
Simonsson, M., Bergholm, J., Lemarchand, D., and Hillier, S.: Mineralogy and biogeochemistry of potassium in the Skogaby experimental forest, southwest Sweden: pools, fluxes and K/Rb ratios in soil and biomass, Biogeochemistry, 131, 77–102, 2016.
Skinner, M. F., Zabowski, D., Harrison, R., Lowe, A., and Xue, D.: Measuring the cation exchange capacity of forest soils, Commun. Soil Sci. Plant Anal., 32, 1751–1764, https://doi.org/10.1081/CSS-120000247, 2001.
Skyllberg, U., Raulund-Rasmussen, K., and Borggaard, O. K.: pH buffering in acidic soils developed under Picea abies and Quercus robur – effects of soil organic matter, adsorbed cations and soil solution ionic strength, Biogeochemistry, 56, 51–74, 2001.
Snäll, S. and Liljefors, T.: Leachability of major elements from minerals in strong acids, J. Geochem. Explor., 71, 1–12, 2000.
Sparks, D.: Potassium Dynamics in Soils, edited by: Stewart, B. A., Advances in Soil Science, Springer, New York, 1–63, 1987.
Starr, M.: Integrated monitoring in Sweden, AMBIO, 40, 833–934, 2011.
Stendahl, J., Snäll, S., Olsson, M. T., and Holmgren, P.: Influence of soil mineralogy and chemistry on site quality within geological regions in Sweden, Forest Ecol. Manag., 170, 75–88, 2002.
Sutherland, R. A.: Multi-element removal from road-deposited sediments using weak hydrochloric acid, Environ. Geol., 42, 937–944, 2002.
Terrer, C., Vicca, S., Hungate, B. A., Phillips, R. P., and Prentice, I. C.: Mycorrhizal association as a primary control of the CO2 fertilization effect, Science, 353, 72–74, 4610, 2016.
Thiffault, E., Hannam, K. D., Paré, D., Titus, B. D., Hazlett, P. W., Maynard, D. G., and Brais, S.: Effects of forest biomass harvesting on soil productivity in boreal and temperate forests – a review, Environ. Rev., 19, 278–309, 2011.
Tipping, E.: Cation binding by humic substances, Vol. 12, Cambridge University Press, 14 pp., 2002.
Van Breemen, N., Driscoll, C. T., and Mulder, J.: Acidic deposition and internal proton sources in acidification of soils and waters, Nature, 307, 599–604, 1984.
Vance, E. D., Brookes, P. C., and Jenkinson, D. S.: Microbial biomass measurments in forest soils: the use of the chloroform fumigation-incubation method in strongly acid soils, Soil Bio. Biochem., 19, 697–702, 1987.
van der Heijden, G., Legout, A., Pollier, B., Ranger, J., and Dambrine, E.: The dynamics of calcium and magnesium inputs by throughfall in a forest ecosystem on base poor soil are very slow and conservative: evidence from an isotopic tracing experiment (26Mg and 44Ca), Biogeochemistry, 118, 413–442, 2014.
Vangansbeke, P., De Schrijver, A., De Frenne, P., Verstraeten, A., Gorissen, L., and Verheyen, K.: Strong negative impacts of whole tree harvesting in pine stands on poor, sandy soils: A long-term nutrient budget modelling approach, Forest Ecol. Manag., 356, 101–111, 2015.
Vuorenmaa, J., Augustaitis, A., Beudert, B., Bochenek, W., Clarke, N., de Wit, H.A., Dirnböck, T., Frey, J., Hakola, H., Kleemola, S., Kobler, J., Krám, P., Lindroos, A.-J., Lundin, L., Löfgren, S., Marchetto, A., Pecka, T., Schulte-Bisping, H., Skotak, K., Srybny, A., Szpikowski, J., Ukonmaanaho, L., Váňa, M., Åkerblom, S., and Forsius, M.: Long-term changes (1990–2015) in the atmospheric deposition and runoff water chemistry of sulphate, inorganic nitrogen and acidity for forested catchments in Europe in relation to changes in emissions and hydrometeorological conditions, Sci. Total Environ., 625, 1129–1145, 2018.
Warfvinge, P. and Sverdrup, H.: Calculating critical loads of acid deposition with PROFILE: a steady-state soil chemistry model, Water Air Soil Pollut., 63, 119–143, 1992.
Yamashita, K., Honjo, H., Nishida, M., Kimura, M., and Asakawa, S.: Estimation of microbial biomass potassium in paddy field soil, Soil Sci. Plant Nutr., 60, 512–519, https://doi.org/10.1080/00380768.2014.919237, 2014.
Zetterberg, T., Köhler, S. J., and Löfgren, S.: Sensitivity analyses of MAGIC modelled predictions of future impacts of whole-tree harvest on soil calcium supply and stream acid neutralizing capacity, Sci. Total Environ., 494/495, 187–201, 2014.
Zetterberg, T., Olsson, B. A., Löfgren, S., Hyvönen, R., and Brandtberg, P. O.: Long-term soil calcium depletion after conventional and whole-tree harvest, Forest Ecol. Manag., 369, 102–115, 2016.
- Abstract
- Introduction
- Methods
- Results and discussion
- Importance of non-exchangeable or acid-extractable base cation pools to nutrient balance concerns in forests
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Special issue statement
- Acknowledgements
- Financial support
- Review statement
- References
- Supplement
- Abstract
- Introduction
- Methods
- Results and discussion
- Importance of non-exchangeable or acid-extractable base cation pools to nutrient balance concerns in forests
- Conclusions
- Data availability
- Author contributions
- Competing interests
- Special issue statement
- Acknowledgements
- Financial support
- Review statement
- References
- Supplement