the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 16 Jun 2025
| 16 Jun 2025
Methane oxidation potential of soils in a rubber plantation in Thailand affected by fertilization
Jun Murase
Kannika Sajjaphan
Chatprawee Dechjiraratthanasiri
Ornuma Duangngam
Rawiwan Chotiphan
Wutthida Rattanapichai
Wakana Azuma
Makoto Shibata
Poonpipope Kasemsap
Daniel Epron
Forest soils, as crucial sinks for atmospheric methane in terrestrial ecosystems, are significantly impacted by changes in ecosystem dynamics due to deforestation and agricultural practices. This study investigated the methane oxidation potential of rubber plantation soils in Thailand, focusing on the effect of fertilization. The methane oxidation activity of the topsoils (0–10 cm) in the dry season was extremely low and increased slightly in the wet season, with lower activity for higher fertilization levels. The methane oxidation potential of the topsoil was too low to explain the in situ methane uptake. Soils below 10 cm depth in unfertilized rubber plantations showed higher activity than the surface soils, and methane oxidation was detected down to, at least, 60 cm depth. In contrast, soils under the high-fertilization treatment exhibited similarly low activity of methane oxidation up to 60 cm depth compared to surface soils during both dry and wet seasons, indicating that fertilization of para rubber plantations negatively impacts the methane oxidation potential of the soils over the deep profile without recovery in the dry (off-harvesting) season with no fertilization. Methane uptake per area, estimated by integrating the methane oxidation potentials of soil layers, was comparable to the field flux data, suggesting that methane oxidation in the soil predominantly occurs at depths below the surface layer. These findings have significant implications for understanding the environmental impacts of tropical forest land uses on methane dynamics and underscore the importance of understanding methane oxidation processes in soils.
- Article
(1600 KB) - Full-text XML
-
Supplement
(586 KB) - BibTeX
- EndNote




Methane is the most important anthropogenically enhanced greenhouse gas in the atmosphere after CO2 (Forster et al., 2021). It is thus important to fully quantify and characterize all sources and sinks to include the role of terrestrial ecosystems in mediating atmospheric exchange. Unsaturated aerobic soils are important sinks of atmospheric methane via oxidation by methane-oxidizing bacteria, with a global estimation of 11–49 Tg CH4 yr−1 (Saunois et al., 2020). The global mean methane uptake rate in forest soils is reported to be 3.95 ± 1.78 kg CH4 ha−1 yr−1, with a total sink of 14.98 ± 6.75 Tg CH4 yr−1 in 1999–2020, thus playing an essential role in the terrestrial methane sink (Feng et al., 2023). Temperate and tropical forest soils are the predominant sinks, contributing 84 % of the total methane sink in forest ecosystems (Feng et al., 2023).
Forest conversion is suspected of weakening the sink of atmospheric methane (Verchot et al., 2000). Rubber plantations in tropical regions have been expanding worldwide, particularly in Asia, where lowered ecosystem functioning compared to forests has been demonstrated (Singh et al., 2021). Deforestation and subsequent agricultural use of tropical forests, such as for para rubber and oil palm plantations, tend to decrease the methane sink of soils (Lang et al., 2019, 2020; Aini et al., 2020; Lang et al., 2017; Zhou et al., 2021). The large-scale expansion of rubber plantations in Southeast Asia has decreased methane uptake by soil. However, a mechanistic understanding of the associated processes within the soil profile is still missing (Lang et al., 2020).
Monitoring the surface methane flux has been used to study the methane uptake of soils in tropical forests and the conversion effects of land use on said uptake. Rubber monocultures showed lower rates of methane uptake than natural forests (Werner et al., 2006), which could turn the rubber soils into methane emitters during a certain period of the rainy season (Lang et al., 2019). The water-filled pore space, which is increased by rubber plantation and with rubber age, is correlated with the methane flux, suggesting that soil compaction by agricultural machinery may suppress methanotrophy and promote methanogenesis by reducing gas exchange (Lang et al., 2019). Methane uptake rates in Indonesian tropical forests under conditions of deforestation and agriculture in the form of rubber plantations are negatively affected by the clay content that controls the soil pore space (Ishizuka et al., 2002).
Mineral nitrogen (ammonia and nitrate) is one of the most critical factors controlling aerobic methane oxidation in soil (Bodelier and Laanbroek, 2004; Bodelier, 2011). Meta-analyses demonstrate that adding mineral nitrogen increases methane emissions and significantly reduces methane uptake in most soil ecosystems (Liu and Greaver, 2009). However, the effect of nitrogen on methane uptake by soils can vary, depending on different factors, such as the enrichment level of nitrogen, the ecosystem, the biome, and the duration of fertilization (Aronson and Helliker, 2010). Low rates of nitrogen addition in forests and tree plantation systems occasionally stimulate methane oxidation (Geng et al., 2017; Koehler et al., 2012); the first study to demonstrate the role of soil fertility in methane uptake in a tropical landscape reported nitrogen limitation of soil methane uptake in the converted land use types, including rubber plantations (Hassler et al., 2015). Another study demonstrated no significant contribution of ammonium nitrogen to predicting methane flux (Lang et al., 2019). Determining the effects of fertilizer application to rubber plantations on soil methane oxidation processes is essential in exploring soil management to compromise between natural rubber production and the maintenance of soil ecosystem functioning as a methane sink.
In this study, we measured the potential rates of soil methane oxidation using a microcosm incubation experiment with the hypothesis that land use change and fertilization management would influence methane oxidation in tropical forest soil, focusing on a para rubber plantation. While most studies assume that methane oxidation in forest soils occurs primarily in the surface soil, we also targeted the deeper soil layers. We tested the hypothesis that the influence of top-dressing fertilizers on soil methane oxidation reaches deeper layers of the soil profile.
2.1 Study site
The study mainly targeted a para rubber (Hevea brasiliensis Müll. Arg.) plantation site at the Sithiporn Kridakorn Research Station, Kasetsart University (SKRS; 10°59.3′ N, 99°29.3′ E), Prachuap Khiri Khan Province, located in southern Thailand. The region's climate is tropical monsoon, with mean annual precipitation of 1700 mm (2010–2023), distributed between a wet season extending from May to November and a dry season from December to April. October and November were the wettest months, with over 250 mm of rain per month on average. The main features of the soil in the SKRS were deep soil with a sandy-loam texture, low water retention capacity, poor organic matter content, and low cation exchange capacity due to the dune origin. The soil was classified as Rhodic Kandiudult.
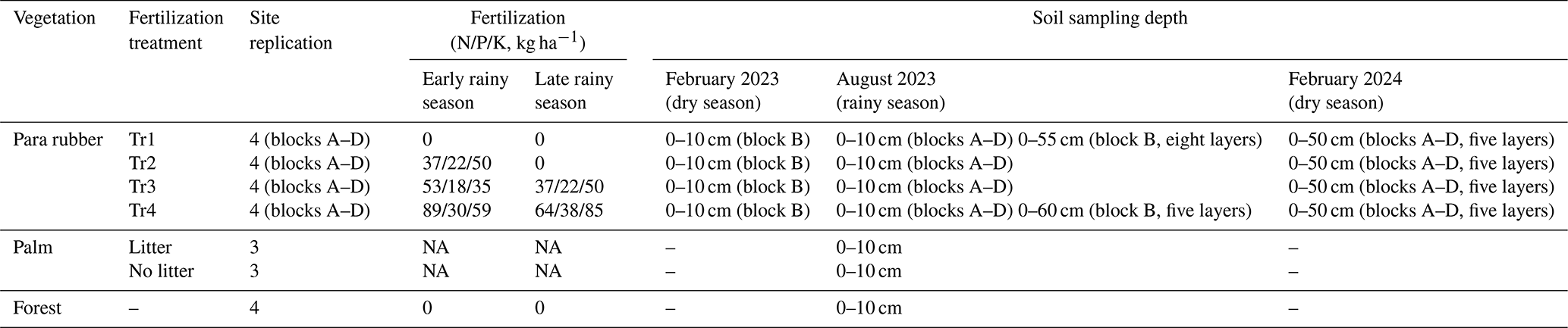
The rubber plantation of the SKRS site was set up in 2007, and four different levels of fertilization have been applied to four randomized replicate blocks (A–D) since 2014 at the beginning of tapping. The four fertilization treatments (Tr) are as follows: Tr1 – no fertilization, Tr2 – low fertilization, Tr3 – intermediate fertilization, and Tr4 – high fertilization (Table 1). Tr3 falls within the range of the fertilizer application rates recommended for mature rubber plantations in Thailand by Thai public institutions, recommendations exceeded by 40 % of rubber farmers (Chambon et al., 2018). A chemical fertilizer composed of nitrogen (40 % nitrate and 60 % ammonium), phosphorus, and potassium (YaraMila™, Yara International ASA, Oslo, Norway) was top-dressed in the wet season, being evenly applied to half of the area between the planting rows. The fertilizer was applied only in the early rainy season (May) for Tr2, while a second application was made late in the rainy season (October) for Tr3 and Tr4. A secondary forest plantation and an oil palm plantation adjoining the rubber plantation in SKRS were also studied for comparison. The bulk density of the soils ranged from 1.4 g cm−3 (forest and palm) to 1.5 g cm−3 (para rubber); the surface of the palm soil covered with palm leaf detritus had a low density (0.94 g cm−3).
Table 1Sampling sites of Sithiporn Kridakorn Research Station. NA – not available.
A rubber plantation at Chachoengsao Rubber Research Center (CRRC; 13°33.9′ N, 101°27.3′ E), Chachoengsao Province, located in central Thailand, was also studied to compare the effect of fertilization on soil methane oxidation with that at SKRS. The annual precipitation was 1400 mm on average for 2022–2023. The soil was classified as clayey skeletal, kaolinitic, and isohyperthermic Typic Kandiustult. Some physicochemical properties of the CRRC soils have been reported before (Kanpanon et al., 2015; Satakhun et al., 2013). The plantation received chemical fertilizer twice yearly at 500 g per tree (N : P : K = ) in the middle of the inter-row, similarly to Tr3 at SKRS.
2.2 Sample collection
Soil samples were collected from the 0–10 cm layer in the middle of the inter-row of the rubber plantation in February 2023 and 2024 (dry season) and August 2023 (wet season). Each month's average temperature was 25.4, 27.2, and 26.5 °C, respectively. In February 2023, triplicate soil samples were collected from one of the replicate blocks (block B). In August 2023 and February 2024, a soil sample was collected from each block, giving four replicate samples per treatment. Soil samples with up to 60 cm depths were also collected at the SKRS rubber plantation site for Tr1 and Tr4 of block B in August 2023 and for all treatments of all replicate blocks in February 2024. Topsoils (0–10 cm) of the SKRS forest and palm plantation and of the CRRC rubber plantation were collected in August 2023. Four replicate sites of the forest were randomly selected, and, for the palm plantation, the two contrasting locations, i.e., with and without palm leaf detritus cover, were selected in triplicate. At CRRC, no fertilization experiment was conducted; thus, we selected locations in the middle of the inter-row for fertilized soils and those in the middle of the row for unfertilized soils as per this site's fertilizer application method. The soil samples were sieved (< 2 mm) on site and stored at room temperature to measure methane oxidation potential within a month. The sieved soil samples for chemical analysis were stored at 4 °C.
2.3 Methane oxidation potential
The potential methane oxidation rates (PMORs) of the sieved soils were determined by means of a microcosm incubation experiment; 10 g of sieved soil was put into 50 mL or 100 mL GC vials (Nichiden-Rika Grass, Kobe, Japan). The vials were capped with butyl rubber stoppers and open-top screw caps and injected with 0.25 or 0.5 mL of 1 % methane to give an initial concentration of 50 ppmv in the headspace containing atmospheric air. The samples were incubated in the dark at 25 °C. Gas samples (0.25 mL) were periodically sampled from the headspace, and methane concentrations were measured with a gas chromatograph with a flame ionization detector (GC-2014, Shimadzu, Kyoto, Japan). The PMORs, expressed in ngCH4 g−1 dry soil h−1, were calculated based on the linear regression of the methane concentration decreasing with incubation time, the volume of the GC vials, and the moisture content of the incubated soils by means of the thermogravimetric method. The PMORs of topsoils (0–10 cm) were also expressed in nmol m−2 s−1 to compare with the in situ soil methane flux, assuming the in situ bulk density of the soil to be 1.5 g cm−3, in accordance with our pilot survey. The potential methane oxidation rate per area (PMORarea, nmol m−2 s−1) was estimated by adding up the methane oxidation rates of different layers.
In the above, l is the soil layer; PMOR (ngCH4 g−1 dry soil h−1) is the potential methane oxidation rate; BD is the bulk density (1.5 g cm−3); Th (cm) is the thickness of the soil layer, multiplied by 10 000 to convert the unit from per square centimeter (cm−2) to per square meter (m−2) and divided by 3600 to convert the unit from per hour (h−1) to per second (s−1); and 16 is the molar mass of methane.
2.4 Soil methane flux
Soil methane fluxes of the SKRS rubber plantation were measured a few days before or after soil sampling on 24 (6 replicates × 4 blocks) PVC collars (20 cm in diameter and 13 cm in height) for each fertilization treatment. Collars were covered with a soil chamber (Li 8100-103, LI-COR; Lincoln, USA). Methane flux was calculated from the rate of change in the methane concentration measured using a trace gas analyzer (Li 7810, LI-COR), as described in Epron et al. (2023).
2.5 Soil chemical analysis
Soil pH was measured in distilled water (soil-to-water ratio of 1:1). Soil electrical conductivity was measured using an electrical conductivity (EC) meter after mixing soil with 5-fold-distilled water. Soil organic carbon was measured using the wet oxidation method (Walkley and Black, 1934), and soil total nitrogen was measured using the Kjeldahl method (Bremner, 1996).
2.6 Statistics
Linear regression analysis was performed using Origin 2024 (OriginLab Corporation, Northampton, USA). Differences in terms of PMORs and soil methane fluxes between treatments were tested using a one-way variance analysis and the Kruskal–Wallis test with the post hoc Tukey's HSD (honestly significant difference) and Dunn–Bonferroni tests, respectively (SPSS for Windows, version 22.0). Principal component analysis was conducted using Primer-e 7 (Plymouth, UK) to detect relationships among the different measures of the topsoil of the para rubber plantation.
3.1 Methane oxidation potential of the surface soils
We first focused on the methane oxidation potentials of the 0–10 cm layer of soil based on previous findings that atmospheric methane uptake in soils is highest in the surface layer (Lang et al., 2020). The soil sampled in the dry season (February 2023) showed minor methane oxidation; only a slight decrease in methane concentration was observed in soils receiving no fertilization (Tr1, Fig. 1a). In the wet season (August 2023), a slight but detectable decrease in methane with time was observed in all treatments compared to during the dry season, and Tr1 showed a more significant decrease in methane than the other treatments (Fig. 1b).
The soils collected in August 2023 from sites other than the SKRS rubber plantation showed methane oxidation activities with more considerable variations (Fig. S1 in the Supplement). The forest soils exhibited a significant spatial variation in methane oxidation. Palm soils showed linear methane consumption, and no significant effect of the litter cover was observed. Methane consumption in CRRC rubber plantation soils sampled from the planting rows was higher than that of those sampled from the middle of the inter-rows where fertilizer was spread.
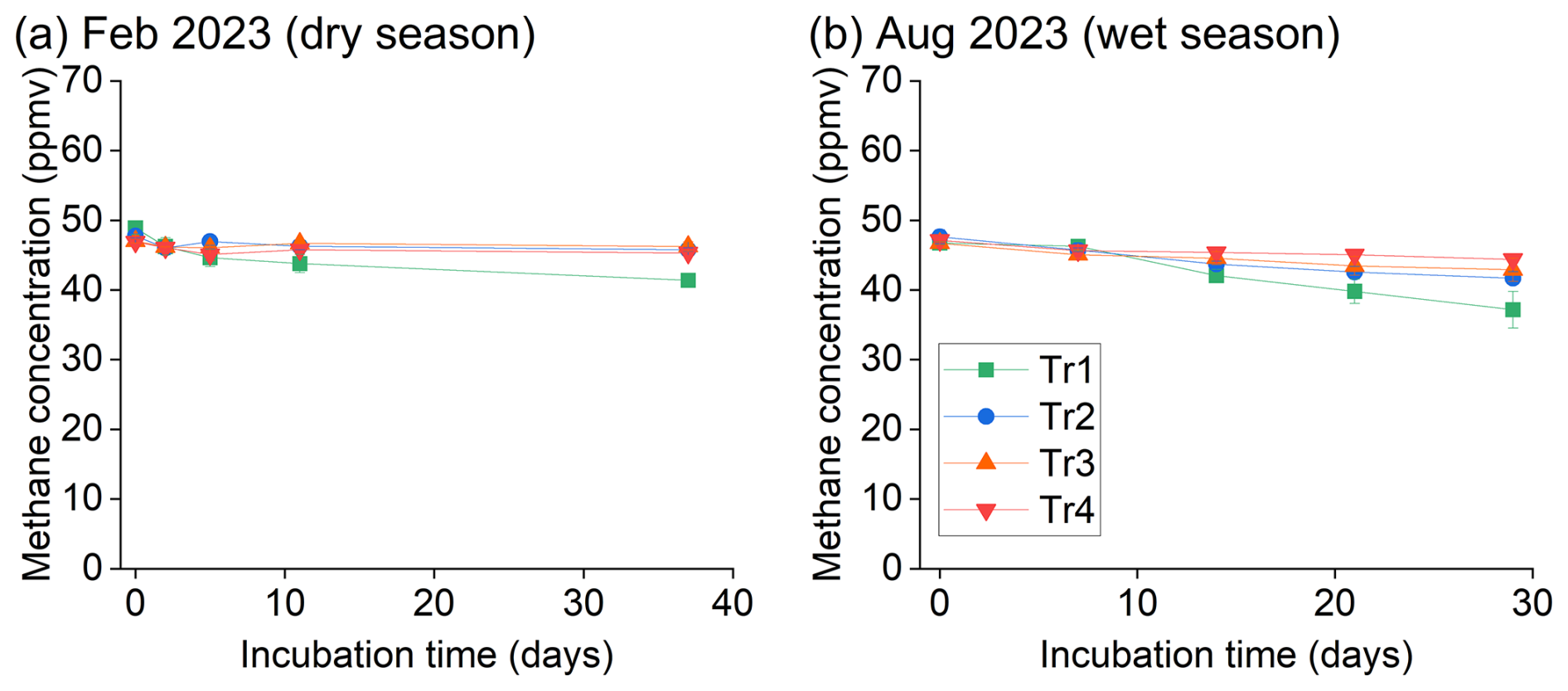
Figure 1Methane oxidation of surface (0–10 cm) soils of the para rubber plantation under different fertilizer treatments in (a) February and (b) August 2023. The February data represent the average and standard error (n=3) of block B, and the August data represent the average and standard error (n=4) of the four blocks (blocks A to D).
The calculated PMORs for the topsoil in August 2023 ranged between 0.02 and 1.23 ngCH4 g−1 dry soil h−1 (Fig. 2). The PMORs of para rubber soils at SKRS ranged from 0.02 to 0.17 ngCH4 g−1 dry soil h−1, with higher rates in Tr1 (no fertilization) than in Tr4 (highest fertilization) (p<0.05), suggesting that fertilization at a critical level lowered soil methane oxidation, which was consistent with previous studies on forest soils (Liu et al., 2024; Aronson and Helliker, 2010). The PMORs of the SKRS rubber soils were among the lowest values reported for European forest soils (Täumer et al., 2021). The rubber soils of CRRC from unfertilized sites (planting rows) also showed higher methane oxidation rates than those from the fertilized sites (inter-rows, p<0.05). The forest soil showed the most considerable spatial variation; two of the four recorded the highest PMORs in this study, which were in the higher range observed in European forest soils (Täumer et al., 2021). The palm soils also showed considerable variation, with no effect of litter cover. The differences in PMORs between the different land uses (forest, rubber, oil palm) were not significant due to the considerable spatial variation in the forest and palm soils, requiring detailed investigation with increasing sample numbers to obtain conclusive results regarding the impact of vegetation change on a soil methane oxidation potential.
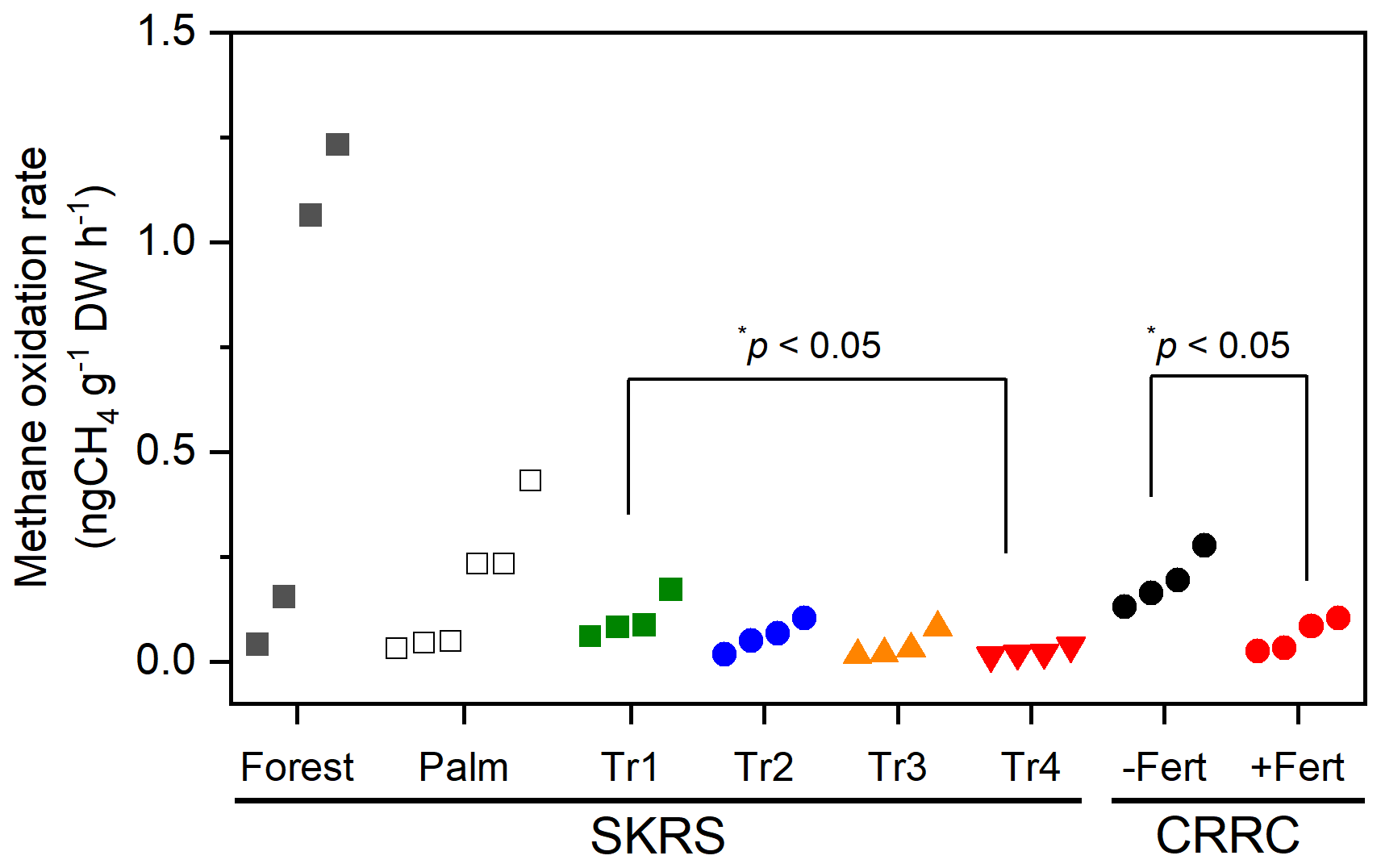
Figure 2Potential methane oxidation rate of the surface (0–10 cm) soils collected in August 2023 (wet season). The unfertilized soils (Tr1 in SKRS and −Fert in CRRC) have higher rates than the fertilized soils (Tr4 and +Fert, respectively).
There was no clear relationship between the PMORs and soil parameters measured in this study, i.e., pH, EC, organic carbon, total nitrogen, and water content (Fig. S2 in the Supplement). A weak positive correlation with total nitrogen was detected, and principal component analysis demonstrated that the PMORs and total nitrogen had similar eigenvalues driven by Tr1 soils, particularly in the wet season (August 2023, Fig. S3). The results may imply that soil organic nitrogen slowly supplies inorganic nitrogen at a rate that does not suppress but rather supports methane-oxidizing bacteria (Geng et al., 2017) in contrast to the high application of chemical fertilizers that often suppress methane oxidation (Liu and Greaver, 2009; Aronson and Helliker, 2010). Water balance is an important factor in regulating the methane dynamics in forest soils (Feng et al., 2020; Bras et al., 2022), but no correlation between soil water content and PMOR was observed in this study. Either drought stress under low soil water content or limited oxygen under high soil water content can inhibit soil methane oxidation (Feng et al., 2020), but soil water contents measured in this study may not have such an inhibitory effect.
The in situ methane flux in the SKRS rubber plantation soil was negative in the dry season (February 2023 and 2024), indicating that methane oxidation predominates methane production, and the soil functioned as a net methane sink. Medium and high fertilization (Tr3 and Tr4) suppressed the in situ soil methane uptake compared to no fertilization (Tr1, Table 2), which was consistent with the results of PMORs. In the wet season (August 2023), the methane fluxes in Tr1 and Tr2 were comparable to those in the dry season, while Tr3 and Tr4 showed positive methane fluxes on average, indicating that methane production in the soil exceeded methane oxidation. The seasonal shift of methane flux from being a sink to a source in a rubber plantation has been reported before (Lang et al., 2019). The estimated aerial PMORs of the surface soil (0–10 cm) were much lower than the methane flux on site in the dry season; the same trend was observed in Tr1 and Tr2 in the wet season. PMORs measured in this study are likely to overestimate the actual oxidation as an initial methane concentration (50 ppmv) higher than the atmospheric level would accelerate methane oxidation (Bender and Conrad, 1994). Thus, the significant gap between the PMORs of topsoil and the methane uptake in situ suggests that the methane oxidation in the topsoil does not explain the in situ methane uptake in the para rubber plantation studied.
Table 2In situ methane flux and PMOR of surface soil (0–10 cm) under different fertilizer treatments in SKRS.
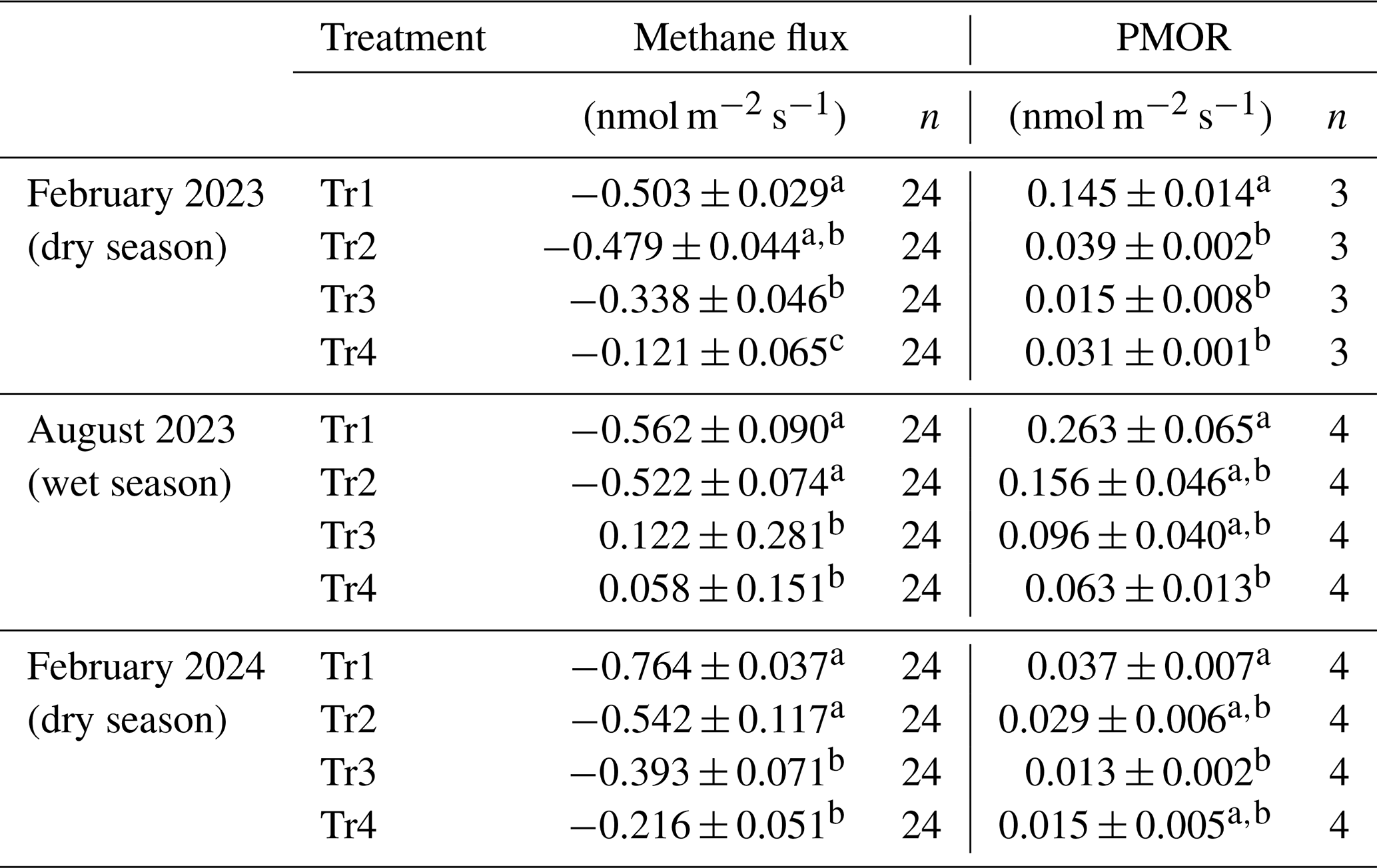
The values are means ± standard errors. The values of different letters are significantly different among the treatments in their respective seasons (P<0.05).
3.2 Methane oxidation potential of the soils collected from deep layers
Subsequently, we tested the hypothesis that the deeper soils could contribute to methane consumption. The soils collected from the layers deeper than 10 cm in the rubber plantation in the dry season (February 2024) showed active methane oxidation compared to the collected topsoil (0–10 cm) in Tr1 (no fertilization) (Figs. 3 and S4); one exception was in block D, where the methane consumption of the topsoil of Tr1 was comparable to that of the deeper layer soils (Fig. S4). Active methane consumption in the deeper soils was also observed in Tr2, except in one replicate block (block A, Fig. S4). Methane oxidation was less active throughout the soil profiles in Tr3 and Tr4 than in Tr1 and Tr2, except in Tr3 in block B, which showed more methane oxidation in the 10–20 cm layer (Fig. 3). Active methane oxidation in the deeper layers under Tr1 and low methane oxidation throughout the soil profiles in Tr4 were also observed in the wet season (Fig. S5).
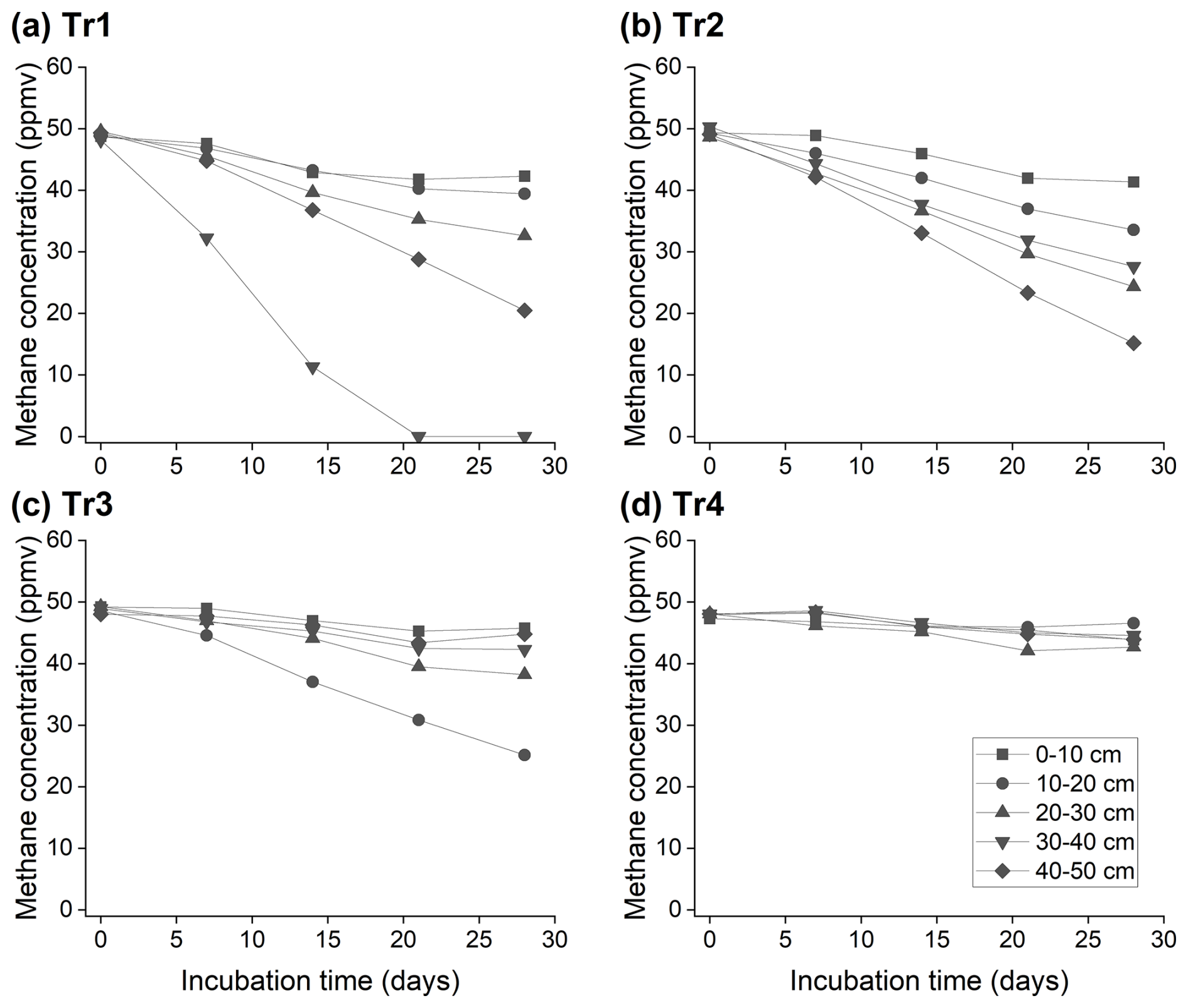
Figure 3Methane oxidation of soils collected from different depths in the rubber plantation (block B) under different fertilizer treatments in February 2024 (dry season).
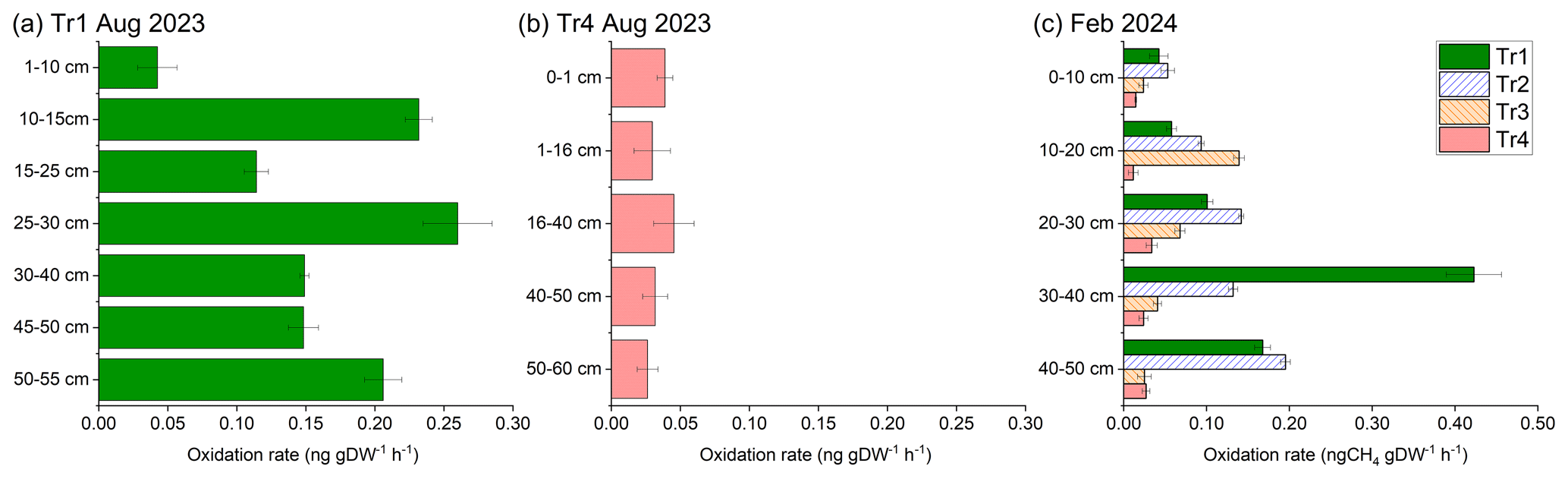
Figure 4Depth profile of potential methane oxidation rate of the SKRS rubber plantation soils (block B) under different fertilizer treatments. The error bars are based on the error of the regression slope. August is in the wet season, and February is in the dry season.
We measured the PMORs of 22 samples from deeper layers (> 10 cm) under Tr1. Among them, 20 samples showed up to 30 times higher PMORs than the topsoil (up to 10 cm) for the same site and sampling time (Figs. 4 and S6). On the other hand, only 3 samples from deeper layers under Tr4, out of 20 samples in total, showed slightly higher PMORs than the topsoil, and the rest of the samples showed PMORs comparable to or lower than the topsoil. Deep soil layers had higher PMORs than the topsoil for 75 % of Tr2 samples (12 out of 16) and 50 % of Tr3 samples (8 out of 16).
The estimated potential methane oxidation rates per area of the SKRS rubber plantation soil tended to decrease along the fertilization level (Fig. 5), and Tr1 and Tr2 showed higher potential methane oxidation rates than Tr3 and Tr4 (p<0.05). Our results demonstrated that (1) unlike the previous studies, the deeper-layer soils in a rubber plantation could contribute to soil methane oxidation, and (2) the top dressing of fertilizers suppressed the soil methane oxidation potentials throughout the soil profile down to at least 60 cm.
In the SKRS rubber plantation, Tr3 and Tr4 soils received more fertilizer than Tr2 soil. In addition, Tr2 soil was fertilized only once a year at the beginning of the wet season, while another fertilizer application was conducted under Tr3 and Tr4 in the late wet season.
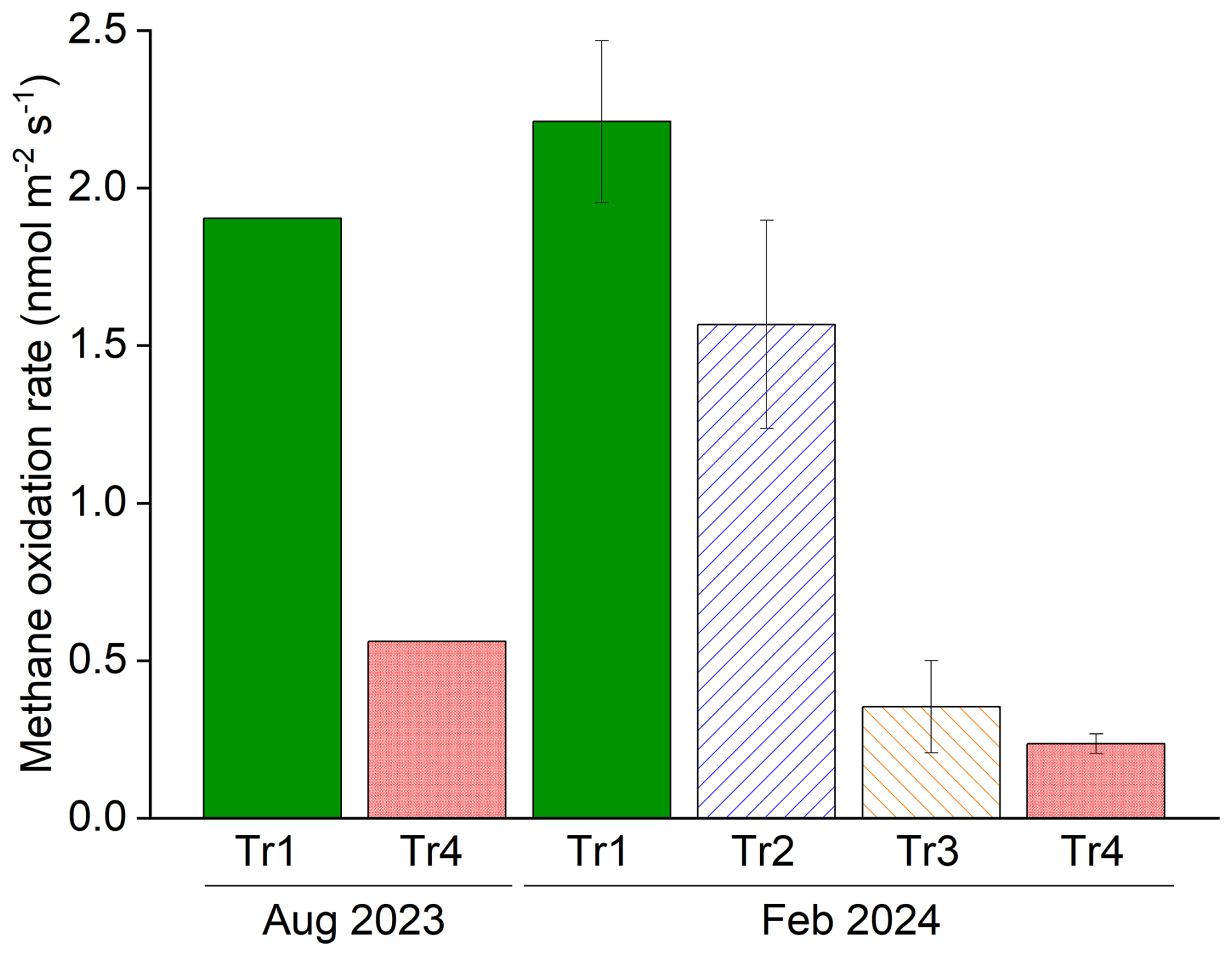
Figure 5Estimated methane oxidation potential per area of SKRS rubber plantation under different fertilizer treatments. Data in August 2023 (wet season) were obtained from block B only, and those in February 2024 (dry season) were the average and standard error of the four replicate blocks (blocks A to D).
Fertilization, especially nitrogen fertilizer application, is often reported to inhibit soil methane oxidation (Täumer et al., 2021; Bodelier, 2011; Bodelier and Laanbroek, 2004). Ammonium competitively suppresses methane monooxygenase due to its similarity to ammonia monooxygenase. Nitrate is also reported to strongly inhibit atmospheric methane oxidation in forest soils (Mochizuki et al., 2012). Both ammonium and nitrate fertilizers are applied to the rubber plantation in this study, which is likely to have suppressed methane oxidation. In addition to the high amount of fertilization, recurring and prolonged disturbances of methane oxidation by fertilization under Tr3 and Tr4 may outcompete the resilience of methane oxidation (Lim et al., 2024). Notably, fertilizers applied on the surface had a suppressive effect on methane oxidation in the deeper layers, at least up to 60 cm. Soil acidification is another possible cause of suppressed methane oxidation in forest soil due to fertilization (Benstead and King, 2001), but there is no relationship between soil pH and potential methane oxidation rates in this study.
The estimated rates ranged between 0.24 (Tr4) and 2.21 nmol m−2 s−1 (Tr1), which exceeded the in situ fluxes. The gaps between in situ methane fluxes and estimated PMORs per area can be related to the fact that the vertical gradients of methane and oxygen concentrations that exist in situ in undisturbed soil profiles were not reproduced in the ex situ incubation in which the soils of each layer were exposed to the same concentrations. Methane oxidation in the soil is insensitive to a wide range of oxygen levels (2 %–20 %) but was suppressed at extremely low oxygen levels (< 2 %) (Walkiewicz and Brzezińska, 2019; Walkiewicz et al., 2018; Bender and Conrad, 1994). Methane concentration in the soil of the study site is often lower than the atmospheric level and is much lower compared to that in the ex situ incubation, even in the hotspots of methane accumulation during the wet season (3.76 ppm) (Epron et al., 2025). PMORs measured for subsoil samples may overestimate the actual oxidation occurring in situ deep in the soil profile (Bender and Conrad, 1994). Another possible explanation is that the in situ fluxes represent net methane uptake, i.e., the balance between oxidation and production, and, thus, could be lower than the gross oxidation rate.
Our findings are in contrast with previous studies that reported higher high- and low-affinity methane oxidation in the topsoil than in the layers below, though some exceptions were noticed when high mineral N concentrations were measured in the topsoil (Reay et al., 2005; Xu et al., 2008). However, in our study, the discrepancy between in situ soil methane uptake and PMORs was observed in all treatments, including Tr1, although the gap was less pronounced in Tr1 than in the three other treatments receiving fertilization. Low soil water content, especially during the dry season, can be another factor pushing methane oxidation down to the soil profile since drought stress is known to inhibit methanotrophic activity (Schnell and King, 1996; Borken et al., 2006; Bras et al., 2022). However, the discrepancy between in situ soil methane uptake and PMORs was observed in all seasons at our site. Alternatively, methane oxidation can be inhibited by several chemical compounds, such as monoterpenes and ethylene, that can be abundant in the upper soil layer under several types of vegetation (Amaral and Knowles, 1998; Jäckel et al., 2004; Maurer et al., 2008). While we did not assess the presence of potential inhibitors of methane oxidation in our study, this hypothesis cannot be ruled out.
The present study demonstrates that high fertilization negatively impacts the methane oxidation potential of soils in the para rubber plantation. The top dressing of fertilizer suppressed methane oxidation not only in the topsoils but also in the deeper-layer soils, which may significantly contribute to the methane cycle in the soil column. In this study, we adapted the sampling strategy over time since the topsoil we collected in our first sampling showed an unexpectedly low methane oxidation potential, unlike in previous studies. We, therefore, subsequently targeted the deeper soil layers. A more systematic study in which high-affinity methane oxidation and methane production are addressed should be carried out in the future. The increase in methane oxidation with depth can be related to a shift in the composition of the methanotrophic community from high- to low-affinity methanotrophs, which remains to be investigated. Nevertheless, our results provide new insights into the impact of tropical agricultural land use on the ecological function of soils in the cycle of a potent greenhouse gas. Even a recommended fertilizer application rate in a rubber plantation could hurt soil methane oxidation potential spatially and temporally, altering the methane cycle of tropical soils. In the future, harmonious land use of tropical soils for rubber plantations should consider the risk of reduced methane uptake due to fertilization. As soil organic nitrogen was weakly but positively correlated with soil methane oxidation potential, soil enrichment with organic nitrogen, e.g., by organic fertilizer application, may be an option to minimize or even reverse the negative impact of fertilization on the methane oxidation of tropical soils, which should be a target of future research.
The data presented in this study are available in the Supplement as an Excel file.
The supplement related to this article is available online at https://doi.org/10.5194/soil-11-457-2025-supplement.
JM: conceptualization (lead), investigation (lead), methodology (lead), supervision (lead), formal analysis (lead), writing (original draft – lead; review and editing – equal). KS: investigation (equal), methodology (equal), writing (review and editing – equal). CD: investigation (equal), methodology (equal), writing (review and editing – equal). OD: investigation (support), writing (review and editing – equal). RC: resources (equal), investigation (support), writing (review and editing – equal). WR: investigation (support), writing (review and editing — equal). WA: investigation (support), writing (review and editing – equal). MS: investigation (support), writing (review and editing – equal). PK: investigation (support), writing (review and editing – equal). DE: conceptualization (support), investigation (equal), writing (review and editing – equal).
The contact author has declared that none of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
This research acknowledges the KAKENHI Grant-in-Aid for Fund for the Promotion of Joint International Research (Fostering Joint International Research(B)) to Daniel Epron. This work was also financially supported by the Office of the Ministry of Higher Education, Science, Research and Innovation and the Thailand Science Research and Innovation through the Kasetsart University Reinventing University program 2023. The authors are grateful to the staff of SKRS and CRRC for their help in the field survey.
This research has been supported by the Japan Society for the Promotion of Science (grant no. 21KK0114, 24KK0127).
This paper was edited by Emily Solly and reviewed by two anonymous referees.
Aini, F. K., Hergoualc'h, K., Smith, J. U., Verchot, L., and Martius, C.: How does replacing natural forests with rubber and oil palm plantations affect soil respiration and methane fluxes?, Ecosphere, 11, e03284, https://doi.org/10.1002/ecs2.3284, 2020.
Amaral, J. A. and Knowles, R.: Inhibition of methane consumption in forest soils by monoterpenes, J. Chem. Ecol., 24, 723–734, https://doi.org/10.1023/a:1022398404448, 1998.
Aronson, E. L. and Helliker, B. R.: Methane flux in non-wetland soils in response to nitrogen addition: a meta-analysis, Ecology, 91, 3242–3251, https://doi.org/10.1890/09-2185.1, 2010.
Bender, M. and Conrad, R.: Methane oxidation activity in various soils and freshwater sediments: Occurrence, characteristics, vertical profiles, and distribution on grain size fractions, J. Geophys. Res.-Atmos., 99, 16531–16540, 1994.
Benstead, J. and King, G. M.: The effect of soil acidification on atmospheric methane uptake by a Maine forest soil1, FEMS Microbiol. Ecol., 34, 207–212, https://doi.org/10.1111/j.1574-6941.2001.tb00771.x, 2001.
Bodelier, P. L. E.: Interactions between nitrogenous fertilizers and methane cycling in wetland and upland soils, Curr. Opin. Environ. Sustain., 3, 379–388, https://doi.org/10.1016/j.cosust.2011.06.002, 2011.
Bodelier, P. L. E. and Laanbroek, H. J.: Nitrogen as a regulatory factor of methane oxidation in soils and sediments, FEMS Microbiol. Ecol., 47, 265–277, https://doi.org/10.1016/s0168-6496(03)00304-0, 2004.
Borken, W., Davidson, E. A., Savage, K., Sundquist, E. T., and Steudler, P.: Effect of summer throughfall exclusion, summer drought, and winter snow cover on methane fluxes in a temperate forest soil, Soil Biol. Biochem., 38, 1388–1395, https://doi.org/10.1016/j.soilbio.2005.10.011, 2006.
Bras, N., Plain, C., and Epron, D.: Potential soil methane oxidation in naturally regenerated oak-dominated temperate deciduous forest stands responds to soil water status regardless of their age – an intact core incubation study, Ann. For. Sci., 79, 29, https://doi.org/10.1186/s13595-022-01145-9, 2022.
Bremner, J. M.: Nitrogen-Total, in: Methods of Soil Analysis, SSSA Book Series, edited by: Sparks, D. L., Page, A. L., Helmke, P. A., Loeppert, R. H., Soltanpour, P. N., Tabatabai, M. A., Johnston, C. T., and Sumner, M. E., Soil Science Society of America, Madison, Wisconsin, USA, 1085–1121,https://doi.org/10.2136/sssabookser5.3.c37, 1996.
Chambon, B., Dao, X. L., Tongkaemkaew, U., and Gay, F.: What determine smallholders' fertilization practices during the mature period of rubber plantations in Thailand?, Exp. Agric., 54, 824–841, https://doi.org/10.1017/S0014479717000400, 2018.
Epron, D., Mochidome, T., Tanabe, T., Dannoura, M., and Sakabe, A.: Variability in stem methane emissions and wood methane production of different tree species in a cold temperate mountain forest, Ecosystems, 26, 784–799, https://doi.org/10.1007/s10021-022-00795-0, 2023.
Epron, D., Chotiphan, R., Wang, Z., Duangngam, O., Shibata, M., Paul, S. K., Mochidome, T., Sathornkich, J., Azuma, W. A., Murase, J., Nouvellon, Y., Kasemsap, P., and Sajjaphan, K.: Fertilization turns a rubber plantation from sink to methane source, EGUsphere [preprint], https://doi.org/10.5194/egusphere-2025-2, 2025.
Feng, H., Guo, J., Peng, C., Ma, X., Kneeshaw, D., Chen, H., Liu, Q., Liu, M., Hu, C., and Wang, W.: Global estimates of forest soil methane flux identify a temperate and tropical forest methane sink, Geoderma, 429, 116239, https://doi.org/10.1016/j.geoderma.2022.116239, 2023.
Feng, H. L., Guo, J. H., Han, M. H., Wang, W. F., Peng, C. H., Jin, J. X., Song, X. Z., and Yu, S. Q.: A review of the mechanisms and controlling factors of methane dynamics in forest ecosystems, Forest Ecol. Manag., 455, 117702, https://doi.org/10.1016/j.foreco.2019.117702, 2020.
Forster, P., Storelvmo, T., Armour, K., Collins, W., Dufresne, J.-L., Frame, D., Lunt, D., Mauritsen, T., Palmer, M., Watanabe, M., Wild, M., and Zhang, H.: Chapter 7: The Earth's energy budget, climate feedbacks, and climate sensitivity, in: Climate Change 2021: The Physical Science Basis, Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, https://doi.org/10.25455/wgtn.16869671, 2021.
Geng, J., Cheng, S., Fang, H., Yu, G., Li, X., Si, G., He, S., and Yu, G.: Soil nitrate accumulation explains the nonlinear responses of soil CO2 and CH4 fluxes to nitrogen addition in a temperate needle-broadleaved mixed forest, Ecol. Indic., 79, 28–36, https://doi.org/10.1016/j.ecolind.2017.03.054, 2017.
Hassler, E., Corre, M. D., Tjoa, A., Damris, M., Utami, S. R., and Veldkamp, E.: Soil fertility controls soil–atmosphere carbon dioxide and methane fluxes in a tropical landscape converted from lowland forest to rubber and oil palm plantations, Biogeosciences, 12, 5831–5852, https://doi.org/10.5194/bg-12-5831-2015, 2015.
Ishizuka, S., Tsuruta, H., and Murdiyarso, D.: An intensive field study on CO2, CH4, and N2O emissions from soils at four land-use types in Sumatra, Indonesia, Global Biogeochem. Cy., 16, 22-21–22-11, https://doi.org/10.1029/2001GB001614, 2002.
Jäckel, U., Schnell, S., and Conrad, R.: Microbial ethylene production and inhibition of methanotrophic activity in a deciduous forest soil, Soil Biol. Biochem., 36, 835–840, 2004.
Kanpanon, N., Kasemsap, P., Thaler, P., Kositsup, B., Gay, F., Lacote, R., and Epron, D.: Carbon isotope composition of latex does not reflect temporal variations of photosynthetic carbon isotope discrimination in rubber trees (Hevea brasiliensis), Tree Physiol., 35, 1166–1175, https://doi.org/10.1093/treephys/tpv070, 2015.
Koehler, B., Corre, M. D., Steger, K., Well, R., Zehe, E., Sueta, J. P., and Veldkamp, E.: An in-depth look into a tropical lowland forest soil: nitrogen-addition effects on the contents of N2O, CO2 and CH4 and N2O isotopic signatures down to 2-m depth, Biogeochem., 111, 695–713, https://doi.org/10.1007/s10533-012-9711-6, 2012.
Lang, R., Blagodatsky, S., Xu, J., and Cadisch, G.: Seasonal differences in soil respiration and methane uptake in rubber plantation and rainforest, Agr. Ecosy. Environ., 240, 314–328, https://doi.org/10.1016/j.agee.2017.02.032, 2017.
Lang, R., Goldberg, S., Blagodatsky, S., Piepho, H.-P., Harrison, R. D., Xu, J., and Cadisch, G.: Converting forests into rubber plantations weakened the soil CH4 sink in tropical uplands, Land Degrad. Dev., 30, 2311–2322, https://doi.org/10.1002/ldr.3417, 2019.
Lang, R., Goldberg, S. D., Blagodatsky, S., Piepho, H.-P., Hoyt, A. M., Harrison, R. D., Xu, J., and Cadisch, G.: Mechanism of methane uptake in profiles of tropical soils converted from forest to rubber plantations, Soil Biol. Biochem., 145, 107796, https://doi.org/10.1016/j.soilbio.2020.107796, 2020.
Lim, J., Wehmeyer, H., Heffner, T., Aeppli, M., Gu, W., Kim, P. J., Horn, M., and Ho, A.: Resilience of aerobic methanotrophs in soils; spotlight on the methane sink under agriculture, FEMS Microbiol. Ecol., 100, fiae008, https://doi.org/10.1093/femsec/fiae008, 2024.
Liu, C.-A., Nie, Y., Zhang, J.-L., Tang, J.-W., Rao, X., and Siddique, K. H. M.: Response of N, P, and metal ions in deep soil layers to long-term cultivation of rubber and rubber-based agroforestry systems, Sci. Total Environ., 946, 174340, https://doi.org/10.1016/j.scitotenv.2024.174340, 2024.
Liu, L. and Greaver, T. L.: A review of nitrogen enrichment effects on three biogenic GHGs: the CO2 sink may be largely offset by stimulated N2O and CH4 emission, Ecol. Lett., 12, 1103–1117, https://doi.org/10.1111/j.1461-0248.2009.01351.x, 2009.
Maurer, D., Kolb, S., Haumaier, L., and Borken, W.: Inhibition of atmospheric methane oxidation by monoterpenes in Norway spruce and European beech soils, Soil Biol. Biochem., 40, 3014–3020, https://doi.org/10.1016/j.soilbio.2008.08.023, 2008.
Mochizuki, Y., Koba, K., and Yoh, M.: Strong inhibitory effect of nitrate on atmospheric methane oxidation in forest soils, Soil Biol. Biochem., 50, 164–166, https://doi.org/10.1016/j.soilbio.2012.03.013, 2012.
Reay, D., Nedwell, D., McNamara, N., and Ineson, P.: Effect of tree species on methane and ammonium oxidation capacity in forest soils, Soil Biol. Biochem., 37, 719–730, https://doi.org/10.1016/j.soilbio.2004.10.004, 2005.
Satakhun, D., Gay, F., Chairungsee, N., Kasemsap, P., Chantuma, P., Thanisawanyangkura, S., Thaler, P., and Epron, D.: Soil CO2 efflux and soil carbon balance of a tropical rubber plantation, Ecol. Res., 28, 969–979, https://doi.org/10.1007/s11284-013-1079-0, 2013.
Saunois, M., Stavert, A. R., Poulter, B., Bousquet, P., Canadell, J. G., Jackson, R. B., Raymond, P. A., Dlugokencky, E. J., Houweling, S., Patra, P. K., Ciais, P., Arora, V. K., Bastviken, D., Bergamaschi, P., Blake, D. R., Brailsford, G., Bruhwiler, L., Carlson, K. M., Carrol, M., Castaldi, S., Chandra, N., Crevoisier, C., Crill, P. M., Covey, K., Curry, C. L., Etiope, G., Frankenberg, C., Gedney, N., Hegglin, M. I., Höglund-Isaksson, L., Hugelius, G., Ishizawa, M., Ito, A., Janssens-Maenhout, G., Jensen, K. M., Joos, F., Kleinen, T., Krummel, P. B., Langenfelds, R. L., Laruelle, G. G., Liu, L., Machida, T., Maksyutov, S., McDonald, K. C., McNorton, J., Miller, P. A., Melton, J. R., Morino, I., Müller, J., Murguia-Flores, F., Naik, V., Niwa, Y., Noce, S., O'Doherty, S., Parker, R. J., Peng, C., Peng, S., Peters, G. P., Prigent, C., Prinn, R., Ramonet, M., Regnier, P., Riley, W. J., Rosentreter, J. A., Segers, A., Simpson, I. J., Shi, H., Smith, S. J., Steele, L. P., Thornton, B. F., Tian, H., Tohjima, Y., Tubiello, F. N., Tsuruta, A., Viovy, N., Voulgarakis, A., Weber, T. S., van Weele, M., van der Werf, G. R., Weiss, R. F., Worthy, D., Wunch, D., Yin, Y., Yoshida, Y., Zhang, W., Zhang, Z., Zhao, Y., Zheng, B., Zhu, Q., Zhu, Q., and Zhuang, Q.: The Global Methane Budget 2000–2017, Earth Syst. Sci. Data, 12, 1561–1623, https://doi.org/10.5194/essd-12-1561-2020, 2020.
Schnell, S. and King, G. M.: Responses of methanotrophic activity in soils and cultures to water stress, Appl. Environ. Microbiol., 62, 3203–3209, 1996.
Singh, A. K., Liu, W. J., Zakari, S., Wu, J. E., Yang, B., Jiang, X. J., Zhu, X. A., Zou, X., Zhang, W. J., Chen, C. F., Singh, R., and Nath, A. J.: A global review of rubber plantations: Impacts on ecosystem functions, mitigations, future directions, and policies for sustainable cultivation, Sci. Total Environ., 796, 148948, https://doi.org/10.1016/j.scitotenv.2021.148948, 2021.
Täumer, J., Kolb, S., Boeddinghaus, R. S., Wang, H., Schöning, I., Schrumpf, M., Urich, T., and Marhan, S.: Divergent drivers of the microbial methane sink in temperate forest and grassland soils, Glob. Change Biol., 27, 929–940, https://doi.org/10.1111/gcb.15430, 2021.
Verchot, L. V., Davidson, E. A., Cattânio, J. H., and Ackerman, I. L.: Land-use change and biogeochemical controls of methane fluxes in soils of eastern Amazonia, Ecosystems, 3, 41–56, https://doi.org/10.1007/s100210000009, 2000.
Walkiewicz, A. and Brzezińska, M.: Interactive effects of nitrate and oxygen on methane oxidation in three different soils, Soil Biol. Biochem., 133, 116–118, https://doi.org/10.1016/j.soilbio.2019.03.001, 2019.
Walkiewicz, A., Brzezińska, M., and Bieganowski, A.: Methanotrophs are favored under hypoxia in ammonium-fertilized soils, Biol. Fert. Soils, 54, 861–870, https://doi.org/10.1007/s00374-018-1302-9, 2018.
Walkley, A. J. and Black, I. A.: An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method, Soil Sci., 37, 29–38, https://doi.org/10.1097/00010694-193401000-00003, 1934.
Werner, C., Zheng, X., Tang, J., Xie, B., Liu, C., Kiese, R., and Butterbach-Bahl, K.: N2O, CH4 and CO2 emissions from seasonal tropical rainforests and a rubber plantation in Southwest China, Plant Soil, 289, 335–353, https://doi.org/10.1007/s11104-006-9143-y, 2006.
Xu, X., Yuan, B., and Wei, J.: Vertical distribution and interaction of ethylene and methane in temperate volcanic forest soils, Geoderma, 145, 231–237, https://doi.org/10.1016/j.geoderma.2008.03.010, 2008.
Zhou, W., Zhu, J., Ji, H., Grace, J., Sha, L., Song, Q., Liu, Y., Bai, X., Lin, Y., Gao, J., Fei, X., Zhou, R., Tang, J., Deng, X., Yu, G., Zhang, J., Zheng, X., Zhao, J., and Zhang, Y.: Drivers of difference in CO2 and CH4 emissions between rubber plantation and tropical rainforest soils, Agr. Forest Meteorol., 304–305, 108391, https://doi.org/10.1016/j.agrformet.2021.108391, 2021.