the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 02 Oct 2025
| 02 Oct 2025
What if publication bias is the rule and net carbon loss from priming the exception?
Jennifer Michel
Yves Brostaux
Bernard Longdoz
Hervé Vanderschuren
Pierre Delaplace
Priming effects in soil science describe the influence of fresh carbon (C) inputs on rates of microbial mineralisation of native soil organic matter, which can either increase (positive priming) or decrease (negative priming). While both positive and negative priming effects occur in natural ecosystems, the latter is less documented in the peer-reviewed literature and the overall impact of priming effects on the C balance of vegetated ecosystems remains elusive. Here, we highlight three aspects which need to be discussed to ensure (rhizosphere) priming effects are correctly perceived in their ecological context and measured at appropriate scales: (i) We emphasize the importance of evaluating net C balances because usually experimental C inputs exceed C-losses meaning even positive priming doesn't cause net C-loss; (ii) We caution against publication bias, which forces overrepresentation of positive priming effects, neglects negative or no priming, and potentially misguides conclusions about C-loss; and (iii) We highlight the need to distinguish between general priming effects and rhizosphere-specific priming, which differ in their scale and driving factors, and hence require different methodological approaches. Future research should focus on scalable experiments linking priming to plant nutrition via C, nutrient and water cycling to understand priming in context of ecosystem functioning.
- Article
(2098 KB) - Full-text XML
- BibTeX
- EndNote




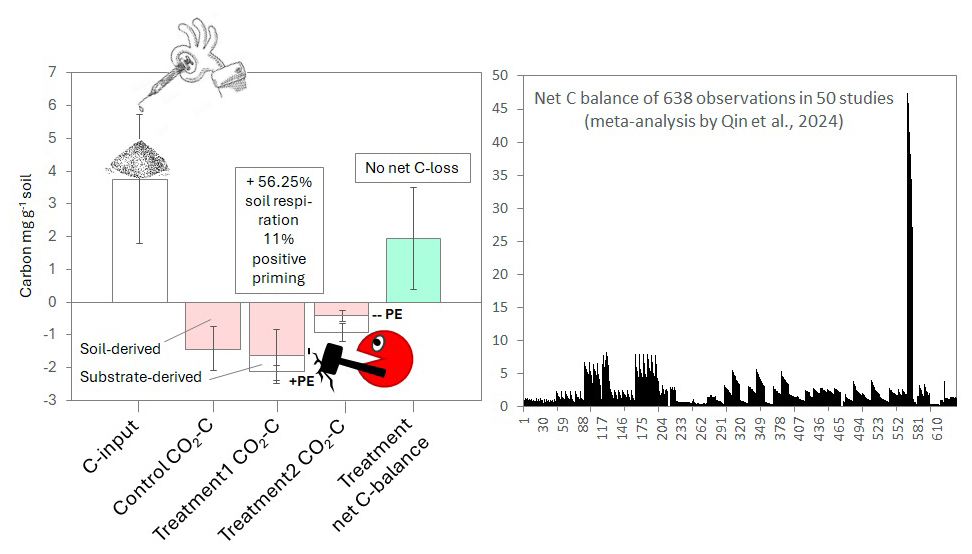
Figure 1Net C balance. Left: Principle of C balance calculation (sum of C inputs minus sum of C output) on a common soil incubation data set with positive (treatment 1) and negative (treatment 2) priming, and no net C loss in neither case because a lot of added C input is not respired and hence stayed in the system either in microbial biomass or dissolved organic C. Right: Net C balance of the n= 638 observations of n= 50 priming studies included in the meta-analysis of Qin et al. (2024).
Priming effects refer to changes in soil C mineralisation rates caused by exogenous C inputs to soil such as litter, while rhizosphere priming effects more specifically refer to the changes in soil microbial activity and nutrient cycling caused by root C inputs to soil by plants. The carbon (C) compounds in root exudates and litter can either stimulate microbial growth and metabolism, leading to increased mineralization of soil organic matter (positive priming), or decrease microbial soil mineralisation when microbes assimilate primarily plant-derived C (negative priming) (Kuzyakov et al., 2000; Blagodatskaya et al., 2011; Dijkstra et al., 2013). Both positive and negative priming effects are commonly reported in the literature, and they are not mutually exclusive in ecosystems (Bastida et al., 2019; Feng et al., 2021; Michel et al., 2024). In many studies, observations include both positive and negative priming either depending on experimental condition, or sometimes substrate amendments also result in mixed positive, negative and/or no priming within one unique modality (Cheng et al., 2014; Qiao et al., 2016; Heitkötter et al., 2017; Hicks et al., 2019; Michel et al., 2022). Individual priming effects are mostly short-term phenomena, but continuously occur in the rhizosphere of living plants, where active root exudation provides energy-rich labile C to soil microbes, while rhizodeposition also supplies more complex substances like cellulose to the soil (Canarini et al., 2019; Villarino et al., 2021). While it is increasingly recognised that priming effects are an important mechanism to regulate plant nutrition, the impact of priming effects on the overall C balance remains controversial (Dijkstra et al., 2013; Zhu et al., 2014; Holz et al., 2023, Pausch et al., 2024). C inputs to soil can also directly interact with the abiotic soil matrix and C inputs can also enter the soil food web bypassing microbes, which can cause temporal and spatial shifts in the priming response to fresh inputs (Barreto et al., 2024; Sokol et al., 2024; van Bommel et al., 2024). Here, we highlight three aspects which need to be discussed to ensure (rhizosphere) priming effects are correctly perceived in their ecological context and measured at appropriate scales to avoid a one-sided narrative distorted towards C loss caused by positive priming.
- i.
The first aspect is that there is little empirical evidence for net C losses from priming as in most studies, including those reporting exclusively positive priming effects, the experimentally added quantities of C to the study system exceed the amounts lost in basal and primed respiration.
- ii.
The second aspect is that publication bias is critical, with studies tending to overrepresent positive priming and inferring C loss without empirical evidence.
- iii.
The third aspect is a lack of distinction between priming effects (PE) and rhizosphere priming effects (RPE) which are measured at different scales, have different drivers and therefore differ in their ecological interpretability.
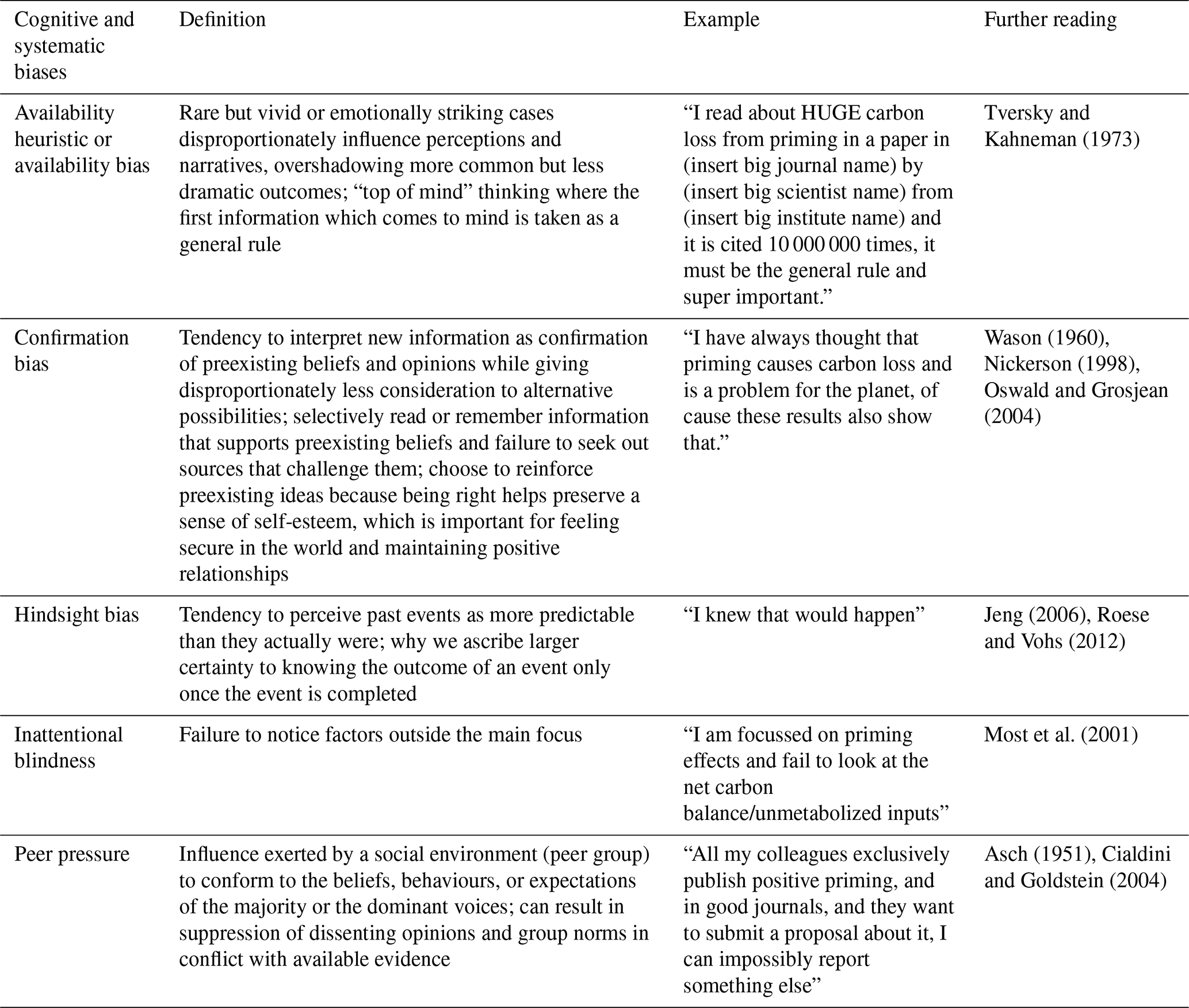
Table 1Cognitive and systemic biases which can influence perception of priming effects (partly after Ruhl, 2023). For an objective analysis free of biases, the essential step is to be aware of the biases (by reading below table e.g.) and engage in discussion of a broader perspective.
1.1 Even positive priming effects seldom cause net carbon loss
Many studies focus on C losses from (positive) priming effects, which has been the historic narrative in priming literature (e.g. Löhnis, 1926; Jenkinson et al., 1985). Positive priming and net C losses are observed in studies, but the number of studies with true C loss is relatively small as commonly the inputs exceed the outputs (Liang et al., 2018). Yet, the small number of studies reporting net C loss and stating huge implications for ecosystem C cycling has a disproportionally strong impact on the overall perception of priming because the results are “catchy”, which can have a strong imprint on the mind (Table 1). Nonetheless, recently more studies provided a more comprehensive view on C budgets and revealed that there is little evidence for net C loss from priming effects (Qiao et al., 2014; Liang et al., 2018; Jian and Bengtson, 2022; Siles et al., 2022; Qin et al., 2024; Chen et al., 2025). For example, a recent meta-analysis evaluating the impact of priming effects derived from crop residues and their interaction with nitrogen inputs concluded that there was no C loss despite the positive priming reported (Qin et al., 2024; Fig. 1). This finding aligns with assessments in many soil incubation studies which demonstrate a net C balance in favour of C sequestration because in these experiments the C inputs from labile substrates usually exceed the C outputs from basal and primed respiration by at least one order of magnitude (Qiao et al., 2014; Cardinael et al., 2015; Liang et al., 2018; Schiedung et al., 2023; Qin et al., 2024). In accordance with these observations in lab incubations, several studies upscaling priming effects over longer time scales and to areas of several hectares also indicate that priming effects may not change overall C budgets. For example, Schiedung et al. (2023) evaluated priming effects along a 20-year chronosequence of land inversion in New Zealand to identify the dependence of priming effects on root-derived C in topsoil and sub soils. Even though positive priming was reported, overall, C losses with priming never exceeded new root-derived C inputs. Similar observations were made by Yin et al. (2019) who studied rhizosphere priming effects and microbial biomass C dynamics of two wheat genotypes grown under two temperatures and found no net soil organic C loss or gain as C loss caused by higher RPE was counteracted by increased microbial growth/turnover. Similarly, Cardinael et al. (2015) used a 52-year long field experiment where SOC stocks of fallow fields were compared to SOC stocks of fields regularly receiving fresh or composted straw to show that no significant difference in SOC stocks dynamics occurred over the 52 years, suggesting no long-term impact of priming effect. Equalising priming with C loss is hence not a valid conclusion and studies should consistently evaluate and present the experimental C inputs and outputs and report the net C balance to avoid misleading the reader to believe that priming imperatively has a strong impact on soil C budgets.
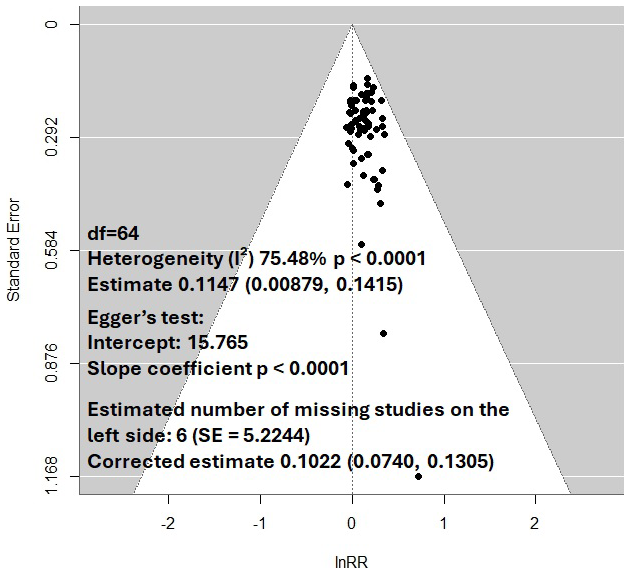
Figure 2Funnel plot after Xu et al. (2024). Funnel plots are evaluated for symmetry: in the absence of bias, they should resemble an inverted funnel, with larger (more precise) studies at the top and smaller (less precise) studies scattered at the base. Asymmetry may suggest publication bias, such as an overrepresentation of small studies with large effects due to selective publication of positive findings. The triangle represents the 95 % confidence interval, and studies outside this interval may indicate heterogeneity (I2) or bias. Heterogeneity reflects inconsistent results caused by variations in study design, populations, interventions, or actual outcomes.
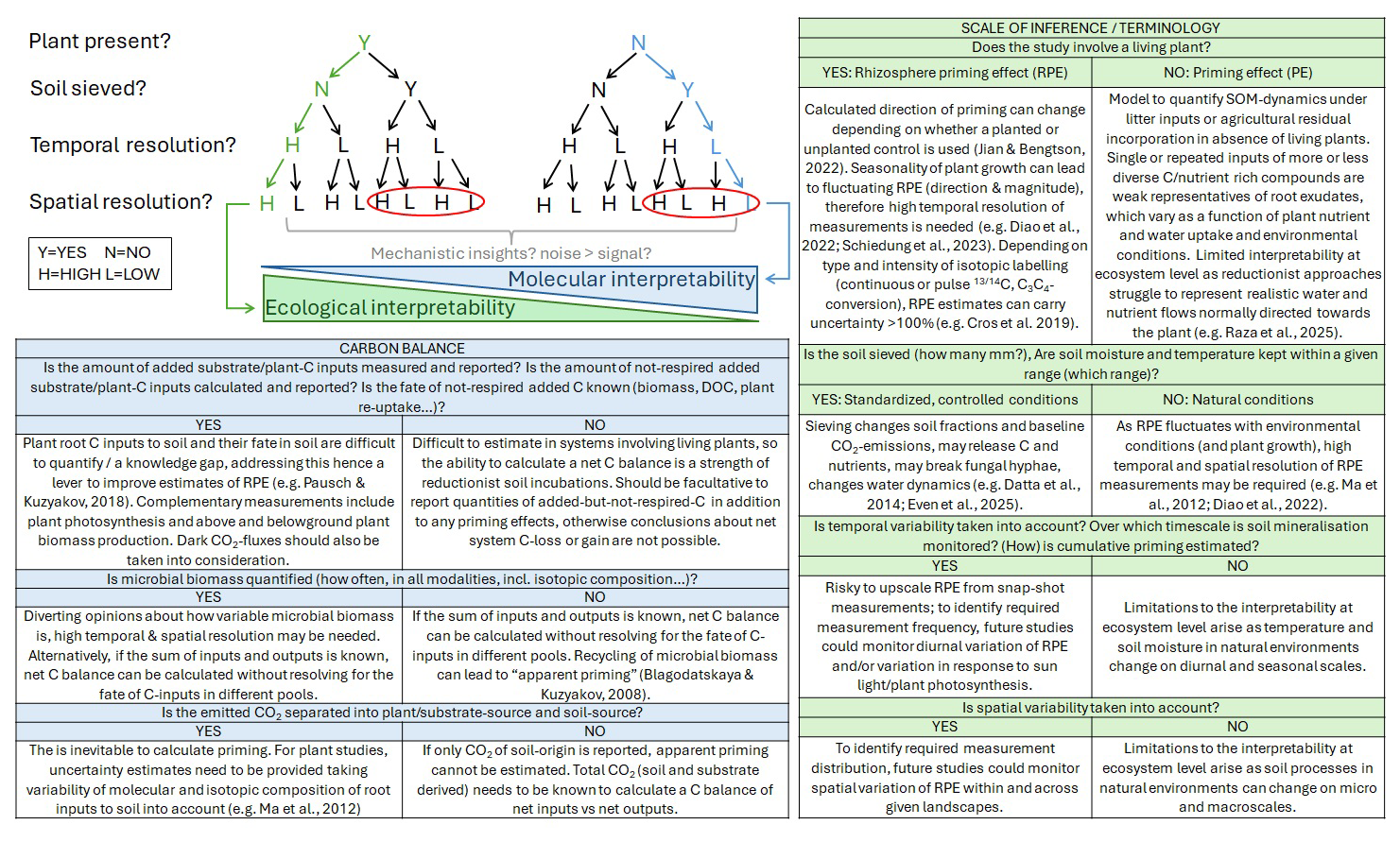
Figure 3Critical checklist to contextualise study design. Red circles indicate common approaches in most experiments. The intermediate paths risk to contain either too much ecological noise to obtain a mechanistic signal, or assume too many simplifications which trigger mechanisms which are rarely to occur in natural terrestrial ecosystems.
1.2 Cognitive and systemic biases cause overrepresentation of positive priming in the literature
The dominance of positive priming in the literature may be inflated by cognitive and systemic biases, which can skew perceptions, research practices, and publication outcomes (Table 1). These biases, including availability heuristic, confirmation bias, hindsight bias, inattentional blindness, and peer pressure, systematically distort the scientific narrative, overemphasizing positive priming while underrepresenting neutral or negative effects. Understanding these biases is critical to foster a balanced scientific discourse and accurately assess the global direction of priming effects. The availability heuristic leads researchers and readers to overestimate the prevalence of positive priming effects due to previous catchy or highly cited studies. For example, a widely publicised study in a prestigious journal claiming dramatic C loss from priming can become “top of mind”, overshadowing more common studies showing minimal or no effects. This bias is compounded by confirmation bias, where researchers may selectively interpret data to align with the prevailing narrative that priming causes significant C loss. For instance, a scientist who believes priming is a major environmental issue might focus on results supporting this view while dismissing contradictory evidence, reinforcing preconceived notions. Hindsight bias further distorts perceptions by making positive priming effects seem more predictable after they are reported. Researchers may claim they knew priming would lead to C loss, even when earlier evidence was ambiguous, solidifying the narrative of positive priming as inevitable. Inattentional blindness contributes by causing researchers to overlook critical factors, such as net C balance or unmetabolized inputs, when focusing narrowly on priming effects. This tunnel vision can lead to incomplete interpretation of data, emphasizing certain outcomes while ignoring broader ecosystem dynamics. Peer pressure plays a significant role in perpetuating such biases, as researchers face social and professional incentives to conform to dominant trends. This systemic pressure contributes to publication bias, where studies reporting positive priming are more likely to be submitted and accepted, while those showing neutral or negative effects are underrepresented, creating an asymmetrical body of literature. In meta-analysis, graphical tools like funnel plots are commonly used to detect publication bias (Fig. 2). These plots display effect sizes (e.g. response ratios) against a measure of study precision (e.g. standard error). Symmetrical plots suggest balanced reporting, while asymmetry – often with a skew toward positive effects – indicates potential bias, where smaller studies with large positive effects are overrepresented. High heterogeneity (e.g. I2>75 %) in these analyses often reflects variability in study methods or selective reporting (aka biases), further complicating the synthesis of global priming effects. Corrective methods in meta-analysis such as trim-and-fill can estimate missing studies to adjust effect sizes (Jennions and Møller, 2002; Cleophas and Zwinderman, 2017; Shi and Lin, 2019). Applying such analysis to the data of a meta-meta-analysis on priming effects by Xu et al. (2024) for example revealed an overall moderate priming estimate of 10.7 % (estimated effect size (log-transformed response ratio) of 0.1022 (CI95: 0.0740, 0.1305)) rather than inflated figures like 125 %). Given that none of the underlying meta-analysis has been corrected for publication bias, the actual priming estimate be even lower. This analysis demonstrating that the interplay of cognitive and systemic biases in scientific literature can strongly distort the representation of priming: When availability heuristic and confirmation bias amplify attention to positive priming, hindsight bias reinforces its perceived inevitability, inattentional blindness narrows focus to supportive data, and peer pressure and publication bias suppress contradictory findings, this can lead to an exaggerated narrative of C loss, potentially misinforming environmental policy and management. To address this, researchers must prioritize transparency, encourage publication of neutral or negative results, and critically evaluate methodological variability (Fig. 3). By mitigating these biases, the scientific community can develop a more accurate and balanced understanding of priming effects and their implications not only for the global C cycle, but also for plant nutrient uptake and the regulation of biogeochemical cycles in natural ecosystems.
1.3 Methodological mismatch? Limited scalability of soil incubations and the need to differentiate priming effects from rhizosphere priming effects
“Priming effects (PE)” refer to interactions between soils, soil microbes and added substances, while “rhizosphere priming effects (RPE)” more specifically describe the interactions between living plant roots, their exudation and other rhizodeposition, rhizosphere microbes and rhizosphere soils. It is important to distinguish between the two, because they differ in their driving factors and the scale of inference (Fig. 3). Priming effects are caused by a static, sometimes repeated, source of substrate input, and usually measured in soil incubation. Rhizosphere priming effects describe changes in SOM mineralisation in the root zone, and are hence subject to dynamic changes in C and nutrient supply and demand, where the plant acts simultaneously as a sink for nutrients and water and a source of C. Hence, several plant physiological parameters like rate of photosynthesis and root exudation are also determinant for rhizosphere priming effects (Dijkstra et al., 2013; Yin et al., 2019; Tang et al., 2019; Raza et al., 2025). It is important to acknowledge the limitations in the scalability of isolated soil incubations to ecosystem processes given that C, nutrient and water pools and fluxes are different in the rhizosphere of living plants as compared to reductionist lab incubations. Moreover, soil incubations are usually conducted under standardised conditions of temperature and soil moisture, and usually soils are sieved before the incubation. Therefore, we have limited knowledge of priming effects in intact soils under variable environmental conditions, and cannot conclude about an impact of priming effects at ecosystem scale based on this data, esp. as the magnitude of priming is usually higher in soil incubations than in the field (Datta et al., 2014; Chen et al., 2023; Even et al., 2025). Hence, it is crucial for future studies to assess whether estimates of priming effect (PE) and mechanistic insights derived from soil incubations accurately reflect processes of rhizosphere priming effects (RPE) in natural ecosystems. Further, more efforts need to be made to measure priming in the field to have a better signal to noise ratio in in-situ studies.
While the soil C priming effect may have limited impact on the C cycle, it is still valuable to evaluate the role of priming in ecosystem functioning, such as how it influences microbial substrate preferences, plant-driven substrate switches, and nutrient dynamics. We suggest a shift from a climate-only perspective to understanding the factors controlling positive and negative priming and their temporal shifts for enhancing plant-soil system resilience and overall ecosystem health. Priming papers should as a rule evaluate the net C balance by juxtapositioning the quantities of primed C and added C to understand whether C has been lost from the system or not. Because often there is no net C loss from soil despite positive priming being reported. To reliably determine the direction of priming across several studies (meta-analysis), publication bias needs to be evaluated very carefully. And prior to that, publication of negative or no priming effects needs to be encouraged. Future studies should also investigate potential discrepancies between soil incubations and field experiments and could address the potential to leverage rhizosphere priming effects to optimise plant nutrition. To upscale (rhizosphere) priming effects to ecosystem processes, their dependency on nutrient, water and temperature dynamics needs to be investigated, which is the opposite of laboratory soil incubations under standardized conditions.
The data presented here is available in the cited papers and respective supplementary materials at https://doi.org/10.1111/gcb.17115 (Qin et al., 2024) and https://doi.org/10.1111/gcb.17502 (Xu et al., 2024).
JM analysed the data and wrote the first draft. All authors critically evaluated the manuscript and approved the final version.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors. Also, please note that this paper has not received English language copy-editing. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
Published with the support of the University Foundation of Belgium.
This paper was edited by Karsten Kalbitz and reviewed by two anonymous referees.
Asch, S. E.: Effects of group pressure upon the modification and distortion of judgments, in: Guetzkow, H., Groups, leadership and men; research in human relations, Carnegie Press, 177–190, https://books.google.be/books?id=DGjQAAAAMAAJ&pg=PA177 (last access: 1 October 2025), 1951.
Barreto, C., Buchkowski, R., and Lindo, Z.: Restructuring of soil food webs reduces carbon storage potential in boreal peatlands, Soil Biol. Biochem., 193, 109413, https://doi.org/10.1016/j.soilbio.2024.109413, 2024.
Blagodatskaya, E. V., Yuyukina, T., Blagodatsky, S., and Kuzyakov, Y.: Turnover of soil organic matter and of microbial biomass under C3–C4 vegetation change: Consideration of 13C fractionation and preferential substrate utilization, Soil Biol. Biochem., 43, 159–166, 2011.
Canarini, A., Kaiser, C., Merchant, A., Richter, A., and Wanek, W.: Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli, Front, Plant Sci., 10, 157, https://doi.org/10.3389/fpls.2019.00157, 2019.
Cardinael, R., Eglin, T., Guenet B., Neill, C., Houot S., and Chenu, C.: Is priming effect a significant process for long-term SOC dynamics? Analysis of a 52-years old experiment, Biogeochemistry, 123, 203–219, https://doi.org/10.1007/s10533-014-0063-2, 2015.
Chen, J., Zhang, Y., Kuzyakov, Y., Wang, D., and Olesen, J. E.: Challenges in upscaling laboratory studies to ecosystems in soil microbiology research, Glob. Change Biol., 29, 569–574, https://doi.org/10.1111/gcb.16537, 2023.
Chen, S., Huang, J., Guo, R., Ma, H., Guo, J., Ling, N., Xu, Q., Wang, M., Shen, Q., and Guo, S.: Soil net carbon balance depends on soil C: N: P stoichiometry, Soil Tillage Res,, 245, 106298, https://doi.org/10.1016/j.still.2024.106298, 2025.
Cheng, W., Parton, W. J., Gonzalez-Meler, M. A., Phillips, R., Asao, S., McNickle, G. G., Brzostek, E. and Jastrow, J. D.: Synthesis and modeling perspectives of rhizosphere priming, New Phytol., 201, 31–44, https://doi.org/10.1111/nph.12440, 2014.
Cialdini, R. B. and Goldstein, N. J.: Social influence: compliance and conformity, Annu. Rev. Psychol., 55, 591–621, https://doi.org/10.1146/annurev.psych.55.090902.142015, 2004.
Cleophas, T. J. and Zwinderman, A. H.: Modern Meta-Analysis. Review and Update of Methodologies, Springer Int. Publ. Switz., https://doi.org/10.1007/978-3-319-55895-0, 2017.
Datta, R., Vranová, V., Pavelka, M., Rejšek, K., and Formánek, P.: Effect of soil sieving on respiration induced by low-molecular-weight substrates, Int. Agrophys., 28, 119–124, https://doi.org/10.2478/intag-2013-0034, 2014.
Dijkstra, F. A., Carrillo, Y., Pendall, E., and Morgan, J. A.: Rhizosphere priming: a nutrient perspective, Front. Microbiol., 4, https://doi.org/10.3389/fmicb.2013.0021, 2013.
Even, R. J., Machmuller, M. B., Lavallee, J. M., Zelikova, T. J., and Cotrufo, M. F.: Large errors in soil carbon measurements attributed to inconsistent sample processing, SOIL, 11, 17–34, https://doi.org/10.5194/soil-11-17-2025, 2025.
Feng, J., Tang, M., and Zhu, B.: Soil priming effect and its responses to nutrient addition along a tropical forest elevation gradient, Glob. Change Biol., 27, 2793–2806, https://doi.org/10.1111/gcb.15587, 2021.
Hicks, L. C., Meir, P., Nottingham, A., Reay, D., Stott, A. W., Salinas, N., and Whitaker, J.: Carbon and nitrogen inputs differentially affect priming of soil organic matter in tropical lowland and montane soils, Soil Biol. Biochem., 129, 212–222, 2019.
Holz, M., Paterson, E., and Pausch, J.: Rhizosphere carbon priming: a plant mechanism to enhance soil nitrogen accessibility?, Plant Soil, 488, 175–185, https://doi.org/10.1007/s11104-023-05979-8, 2023.
Jeng, M. A.: Selected history of expectation bias in physics, Am. J. Phys., 74, 578–583, https://doi.org/10.1119/1.2186333, 2006.
Jenkinson, D. S., Fox, R. H., and Rayner, J. H.: Interactions between fertilizer nitrogen and soil nitrogen – the so-called “priming” effect, J. Soil Sci., 36, 425–444, 1985.
Jennions, M. D. and Møller, A. P.: Publication bias in ecology and evolution: an empirical assessment using the “trim and fill” method, Biol. Rev. Camb. Philos. Soc., 77, 211–222, https://doi.org/10.1017/s1464793101005875, 2002.
Jian, L. I. and Bengtson, P.: Comparative analysis of planted and unplanted controls for assessment of rhizosphere priming effect, Pedosphere, 32, 884–892, 2022.
Kuzyakov, Y., Friedel, J. K., and Stahr, K.: Review of mechanisms and quantification of priming effects, Soil Biol. Biochem., 32, 1485–1498, 2000.
Liang, J., Zhou, Z., Huo, C., Shi, Z., Cole, J. R., Huang, L., Konstantinidis, K. T., Li, X., Liu, B., Luo, Z., Penton, C. R., Schuur, E. A. G., Tiedje, J. M., Wang, Y. P., Wu, L., Xia, J., Zhou, J., and Luo, Y.: More replenishment than priming loss of soil organic carbon with additional carbon input, Nat. Commun., 9, 3175, https://doi.org/10.1038/s41467-018-05667-7, 2018.
Löhnis, F.: Nitrogen availability of green manure, Soil Sci., 22, 253–290, 1926.
Michel, J., Fontaine, S., Revaillot, S., Piccon-Cochard, C., and Whitaker, J.: Plant growth stage and soil horizon respectively determine direction and magnitude of rhizosphere priming effects in contrasting treeline soils, Funct. Ecol., 38, 1859–2080, https://doi.org/10.1111/1365-2435.14625, 2024.
Michel, J., Hartley, I. P., Buckeridge, K. M., van Meegen, C., Broyd, R., Reinelt, L., Ccahuana Quispe, A. J., and Whitaker, J.: Preferential substrate use decreases priming effects in contrasting treeline soils, Biogeochemistry 162, 141–161, https://doi.org/10.1007/s10533-022-00996-8, 2022.
Most, S. B., Simons, D. J., Scholl, B. J., Jimenez, R., Clifford, E., and Chabris, C. F.: How not to be seen: The contribution of similarity and selective ignoring to sustained inattentional blindness, Psychol. Sci., 12, 9–17, 2001.
Nickerson, R. S.: Confirmation bias: A ubiquitous phenomenon in many guises, Rev. Gen. Psychol., 2, 175–220, 1998.
Oswald, M. E. and Grosjean, S.: Confirmation Bias, in: Cognitive Illusions: A Handbook on Fallacies and Biases in Thinking, edited by: Pohl, R. F., Judgement and Memory, Hove, UK: Psychology Press, 79–96, ISBN 978-1-84169-351-4, 2004.
Pausch, J., Holz, M., Zhu, B., and Cheng, W.: Rhizosphere priming promotes plant nitrogen acquisition by microbial necromass recycling, Plant Cell Environ., 1–10, https://doi.org/10.1111/pce.14858, 2024.
Qiao, N., Schaefer, D., Blagodatskaya, E., Zou, X., Xu, X., and Kuzyakov, Y.: Labile carbon retention compensates for CO2 released by priming in forest soils, Glob. Change Biol., 20, 1943–1954, https://doi.org/10.1111/gcb.12458, 2014.
Qiao, N., Xu, X., Hu, Y., Blagodatskaya, E., Liu, Y., Schaefer, D., and Kuzyakov, Y.: Carbon and nitrogen additions induce distinct priming effects along an organic-matter decay continuum, Sci. Rep., 6, 19865, https://doi.org/10.1038/srep19865, 2016.
Qin, J., Chen, N., Scriber, K. E. II, Liu, J., Wang, Z., Yang, K., Yang, H., Liu, F., Ding, Y., Latif, J., and Jia, H.: Carbon emissions and priming effects derived from crop residues and their responses to nitrogen inputs, Glob. Change Biol., 30, e17115, https://doi.org/10.1111/gcb.17115, 2024.
Raza, S., Cooper, H. V., Girkin, N. T., Kent, M. S., Bennett, M. J., Mooney, S. J., and Colombi, T.: Missing the input: the underrepresentation of plant physiology in global soil carbon research, SOIL, 11, 363–369, https://doi.org/10.5194/soil-11-363-2025, 2025.
Roese, N. J. and Vohs, K. D.: Hindsight bias, Perspect. Psychol. Sci., 7, 411–426, 2012.
Ruhl, C.: Cognitive Bias: How We Are Wired to Misjudge, SimplyPsychology via https://www.simplypsychology.org/cognitive-bias.html (last access: 2 July 2025), 2023.
Schiedung, M., Don, A., Beare, M. H., and Abiven, S.: Soil carbon losses due to priming moderated by adaptation and legacy effects, Nat. Geosci., 16, 909–914, https://doi.org/10.1038/s41561-023-01275-3, 2023.
Shi, L. and Lin, L.: The trim-and-fill method for publication bias: practical guidelines and recommendations based on a large database of meta-analyses, Medicine, 98, e15987, https://doi.org/10.1097/MD.0000000000015987, 2019.
Siles, J. A., Díaz-López, M., Vera, A., Eisenhauer, N., Guerra, C. A., Smith, L. C., Buscot, F., Reitz, T., Breitkreuz, C., van den Hoogen, J., Crowther, T. W., Orgiazzi, A., Kuzyakov, Y., Delgado-Baquerizo, M., and Bastida, F.: Priming effects in soils across Europe, Glob. Change Biol., 28, 2146–2157, https://doi.org/10.1111/gcb.16062, 2022.
Sokol, N. W., Foley, M. M., Blazewicz, S. J., Bhattacharyya, A., DiDonato, N., Estera-Molina, K., Firestone, M., Greenlon, A., Hungate, B. A., Kimbrel, J., Liquet, J., Lafler, M., Marple, M., Nico, P. S., Paša-Tolić, L., Slessarev, E., and Pett-Ridge, J.: The path from root input to mineral-associated soil carbon is dictated by habitat-specific microbial traits and soil moisture, Soil Biol. Biochem., 193, 109367, https://doi.org/10.1016/j.soilbio.2024.109367, 2024.
Tang, M., Cheng, W., Zeng, H., and Zhu, B.: Light intensity controls rhizosphere respiration rate and rhizosphere priming effect of soybean and sunflower, Rhizosphere, 9, 97–105, https://doi.org/10.1016/j.rhisph.2018.12.002, 2019.
Tversky, A. and Kahneman, D.: Availability: A heuristic for judging frequency and probability, Cognitive Psychology, 5, 207–232, 1973.
van Bommel, M., Arndt, K., Endress, M. G., Dehghani, F., Wirsching, J., Blagodatskaya, E., Blagodatsky, S., Kandeler, E., Marhan, S., Poll, C., and Ruess, L.: Under the lens: Carbon and energy channels in the soil micro-food web, Soil Biol. Biochem., 199, 109575, https://doi.org/10.1016/j.soilbio.2024.109575, 2024.
Villarino, S. H., Pinto, P., Jackson, R. B., and Piñeiro, G.: Plant rhizodeposition: A key factor for soil organic matter formation in stable fractions, Sci. Adv., 7, eabd3176, https://doi.org/10.1126/sciadv.abd3176, 2021.
Wason, P. C.: On the failure to eliminate hypotheses in a conceptual task, Q. J. Exp. Psychol., 12, 129–40, 1960.
Xu, S., Delgado-Baquerizo, M., Kuzyakov, Y., Wu, Y., Liu, L., Yang, Y., Li, Y., Yu, Y., Zhu, B., and Yao, H.: Positive soil priming effects are the rule at a global scale, Glob. Change Biol., 30, e17502, https://doi.org/10.1111/gcb.17502, 2024.
Yin, L., Corneo, P. E., Richter, A., Wang, P., Cheng, W., and Dijkstra, F. A.: Variation in rhizosphere priming and microbial growth and carbon use efficiency caused by wheat genotypes and temperatures, Soil Biol. Biochem., 134, 54–61, https://doi.org/10.1016/j.soilbio.2019.03.019, 2019.
Zhu, B., Gutknecht, J. L. M., Herman, D. J., Keck, D. C., Firestone, M. K., and Cheng, W.: Rhizosphere priming effects on soil carbon and nitrogen mineralization, Soil Biol. Biochem., 76, 183–192, https://doi.org/10.1016/j.soilbio.2014.04.033, 2014.