the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 08 Jul 2025
| 08 Jul 2025
Evaluating N2O emissions and carbon sequestration in temperate croplands with cover crops: insights from field trials
Victoria Nasser
René Dechow
Mirjam Helfrich
Ana Meijide
Pauline Sophie Rummel
Heinz-Josef Koch
Reiner Ruser
Lisa Essich
Klaus Dittert
Cover crops (CCs) are acclaimed for enhancing the environmental sustainability of agricultural practices by aiding in carbon (C) sequestration and reducing losses of soil mineral nitrogen (SMN) after harvest. Yet, their influence on nitrous oxide (N2O) emissions – a potent greenhouse gas – presents a complex challenge, with findings varying across different studies. This research aimed to elucidate the effects of various winter CCs – winter rye (frost-tolerant grass), saia oat (frost-sensitive grass), and spring vetch (frost-sensitive legume) – compared to a bare fallow control on SMN dynamics, N2O emissions, and C sequestration. These effects were determined by measuring SMN dynamics and N2O emissions in field experiments. The effects of CCs on soil C sequestration over a 50-year period were predicted by soil organic C (SOC) models using measured aboveground and belowground CC biomass. While CCs efficiently lowered SMN levels during their growth, they slightly increased N2O emissions compared to bare fallow. In particular, winter frost events triggered significant emissions from the frost-sensitive varieties. Moreover, residue incorporation and tillage practices were associated with increased N2O emissions in all CC treatments. Winter rye, characterized by its high biomass production and nitrogen (N) uptake, was associated with the highest cumulative N2O emissions, highlighting the influence of biomass management and tillage practices on N cycling and N2O emissions. The CC treatment resulted in a slight increase in direct N2O emissions (4.5±3.0, 2.7±1.4, and for rye, oat, and vetch, respectively) compared to the fallow () over the entire trial period (18 months). However, the potential of non-legume CCs to reduce indirect N2O emissions compared to fallow (0.3±0.4 and for rye and oat, respectively) and their contribution to C sequestration (120–150 over a period of 50 years when CCs were grown every fourth year) might partially counterbalance these emissions. Thus, while CCs provide environmental benefits, their net impact on N2O emissions requires further research into optimized CC selection and management strategies tailored to specific site conditions to fully exploit their environmental advantages.
- Article
(3286 KB) - Full-text XML
-
Supplement
(446 KB) - BibTeX
- EndNote




The use of cover crops (CCs) is currently being strongly promoted in many countries due to the multifaceted agro-ecological benefits they offer. They positively influence soil physical, chemical, and biological properties, enhancing soil water retention and aiding in weed and disease control through competitive interactions and pest cycle disruptions (Adetunji et al., 2020; Araújo et al., 2021). The incorporation of CC residues boosts soil microbial biomass and activity, thereby enriching biodiversity and providing habitats for beneficial insects (Elhakeem et al., 2019; Finney et al., 2017). When managed effectively, CCs can enhance the yield of subsequent main crops (Adetunji et al., 2020). Grunwald et al. (2022) observed that cover cropping prior to sugar beet cultivation improved soil physical properties, facilitating early growth of sugar beet, which is crucial for high sugar yields (Malnou et al., 2006). However, the effect of CCs on soil water storage, succeeding crop yield, and water-use efficiency may not be consistent in all regions (Wang et al., 2021).
Cover crops may also influence nitrous oxide (N2O) emissions, a potent greenhouse gas. The agricultural sector accounts for about 60 % of global anthropogenic N2O emissions (Masson-Delmotte et al., 2021). These emissions have increased since 1980 due to increased nitrogen (N) fertilizer and manure usage (Davidson, 2009; Tian et al., 2020). Approximately half of the N applied to agricultural fields is absorbed by crops, while the remainder is subject to loss into the atmosphere as NH3, NO, N2O, and N2 or loss to groundwater and surface water primarily in the form of nitrate () (Galloway and Cowling, 2002). In temperate regions, N losses are exacerbated during periods of high precipitation (Gabriel and Quemada, 2011). Cover crops can mitigate these losses by absorbing excess soil mineral N (SMN) after harvest (Abdalla et al., 2019), reducing leaching and runoff, and thereby reducing the need for N fertilization in subsequent crops (Constantin et al., 2011; Hanrahan et al., 2021; Nouri et al., 2022; Tonitto et al., 2006). Leguminous CCs further contribute to soil N through atmospheric fixation (Parr et al., 2011). They may also aid in carbon (C) sequestration when used as green manure, enhancing soil C stocks (Poeplau and Don, 2015). Nevertheless, studies on the net effects of CCs on N2O emissions have yielded mixed results (Basche et al., 2014; Guenet et al., 2021).
Cover crop residue management and soil cultivation practices play a crucial role in N2O emission dynamics and magnitudes. Frost-tolerant CCs can be terminated using various methods in preparation for the next cash crop, while frost-sensitive ones are typically terminated by winter frosts (Storr et al., 2021; Wayman et al., 2015). In some cases, greater soil disturbance in conventional tillage was found to increase N2O emissions compared to reduced tillage or no tilling (Chatskikh et al., 2008). On the other hand, plowing of heavy soils was found to significantly reduce N2O emissions (Rochette et al., 2008). Soil incorporation of CC residues increases N2O emissions compared to surface placement, likely due to accelerated decomposition following increased contact with soil microorganisms (Basche et al., 2014; Lynch et al., 2016).
Crop residues are generally seen as contributors to N2O emissions because of their N and C content. According to the IPCC (2019), about 1 % of the N in crop residues is converted to N2O. The biochemical properties of CCs, such as the C : N ratio, are critical in influencing residue decomposition rates and subsequent N2O emissions (Lynch et al., 2016). Residues with lower C : N ratio are decomposed faster, leading to higher N2O emissions (Basche et al., 2014; Chen et al., 2013; Fosu et al., 2007). The developmental stage of CCs at termination is also relevant due to its impact on residue composition and the amount of N2O emissions after incorporation (Balkcom et al., 2015). Early termination of CCs, which are typically characterized by lower C : N ratios, results in higher N2O emissions (Abalos et al., 2022). Legume CCs generally have lower C : N ratios and result in higher N2O emissions than non-legumes (Basche et al., 2014; Muhammad et al., 2019).
While crop residue quality plays the biggest role in predicting crop-residue-induced N2O emissions, environmental factors such as soil pH, soil N, available soil organic C (SOC), water-filled pore space (WFPS), and temperature also play a role (Abalos et al., 2022). In addition, in temperate cold humid zones, freeze–thaw cycles can lead to significant N2O emission peaks, substantially contributing to the annual cropland N2O emissions in these regions (Goodroad and Keeney, 1984; Lemke et al., 1998; Wagner-Riddle et al., 2017).
In numerous meta-studies it has been shown that CCs increase SOC stocks (Abdalla et al., 2019; Blanco-Canqui, 2022; Blanco-Canqui et al., 2015; Bolinder et al., 2020; Poeplau and Don, 2015). Thus, soil C sequestration from CCs needs to be considered when assessing the effect of CCs on greenhouse gas (GHG) emissions from croplands. The magnitude of GHG savings depends on site-specific conditions and additional C inputs from CCs. Carbon turnover models have been applied to model the effects of CCs on SOC sequestration (Poeplau and Don, 2015; Seitz et al., 2023). In these model applications, the modeled effects are sensitive to the additional C inputs from CCs that vary between CC species.
The objective of this study was to assess how CC species – differing in frost tolerance and biochemical composition – influence both short-term and long-term N2O emissions and SMN dynamics in a CC–sugar beet–winter wheat rotation on Luvisol soils, a common substrate for sugar beet cultivation in Germany. We also aimed to identify key drivers of N2O emissions, including soil temperature, moisture, SMN concentrations, and the quantity and composition of incorporated CC biomass, to weigh the benefits and drawbacks of cover cropping with respect to N2O emissions, SMN dynamics, and SOC sequestration.
The following hypotheses were formulated: (i) during the cover-cropping phase, non-legume CCs reduce SMN and subsequent N2O emissions relative to fallow by assimilating excess N in autumn; (ii) freezing of frost-sensitive CCs elevates SMN levels, leading to higher N2O emissions during winter; (iii) CCs with lower C : N ratios, which decompose more rapidly, result in higher N2O emissions after incorporation than those with higher C : N ratios; (iv) incorporation of CCs with greater biomass residues increases N2O emissions in the following main crop; and (v) carbon inputs from aboveground and belowground biomass – and their contributions to SOC stocks – vary significantly among CC species.
2.1 Study sites and experimental design
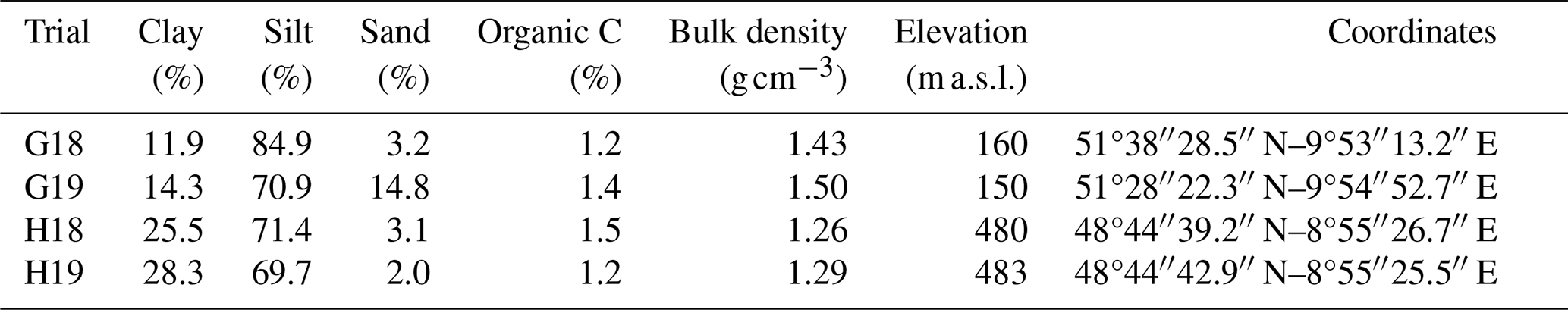
Field trials were conducted in central (Göttingen) and southern Germany (Ihinger Hof, Hohenheim), with replicated fields at each site, established in 2 consecutive years systematically named to reflect their location and establishment year. Specifically, the trials initiated in 2018 were labeled G18 for Göttingen and H18 for Hohenheim, while those initiated in 2019 were labeled G19 and H19, respectively. Each of these trials began in autumn and continued for approximately 18 months, with the 2018 trials ending in March 2020 and the 2019 trials in March 2021. Different fields were used at each site to avoid residual effects between trials. The soils at both sites were classified as Luvisols (IUSS, 2015) with a pH of 7.0–7.5 (in 0.0125 M CaCl2). Details of soil characteristics and site information are presented in Table 1.
Table 1Topsoil properties and site information for the different experimental trials.
Long-term climate data (1991–2020) from the German Meteorological Service (DWD, 2023) recorded average annual precipitation of 624 mm and temperature of 9.4 °C for Göttingen and 701 mm of precipitation with a temperature of 9.1 °C for Hohenheim. Meteorological data, including daily precipitation and hourly soil and air temperatures, were collected from stations located at the field sites.
Field pea (Pisum sativum L.) was cultivated prior to the experiments due to its high residual soil N content and potential for leaching (Voisin et al., 2002). Pea straw was left on the fields and incorporated into the soil by plowing or deep rigid tine cultivator tillage. A randomized complete block design with four replications was set up at each site and in each year. Plot sizes were 21 m×17 m in Göttingen and 30 m×19 m in Hohenheim, with sampling restricted to subplots of 2.7 m×14 m and 3 m×12 m, respectively. Three different CCs were sown in autumn: saia oat (Avena strigosa Schreb. var. “Pratex”) and winter rye (Secale cereale L. var. “Traktor”), representing frost-sensitive and frost-tolerant grasses, respectively, and spring vetch (Vicia sativa L. var. “Mirabella”), a frost-sensitive leguminous CC. These were compared to bare fallow during the CC cultivation period. The management details of the experimental trials are delineated in Table 2. Seedbed preparation was done using a disk or rotary harrow. Seed rates were 120 kg ha−1 for rye, 80 kg ha−1 for oat, and 90 kg ha−1 for vetch, with row spacings of 12.5 cm in Göttingen and 15.0 cm in Hohenheim. During the CC phase, herbicides were applied in autumn for weed control in fallow plots and CC plots were not fertilized. In March of the year following establishment, CCs were treated with glyphosate to terminate any plant growth after winter. To safeguard an optimal seedbed for sugar beet seedlings, rye was plowed to a depth of 30 cm due to its extensive crown root and stem base material, while other treatments were tilled to 15 cm using a short disk harrow or tine cultivator.
Sugar beet (Beta vulgaris L. var. “Lisanna”) followed as the first main crop, sown at 45–50 cm row spacing and 90 000–95 000 plants ha−1 density. In accordance with the German Fertilizer Ordinance (German Fertilizer Ordinance, 2017), the N fertilization for sugar beet was adjusted to a total requirement of 180 kg N ha−1. This total comprised the SMN measured in March in the 0–90 cm horizon and the mineral N applied thereafter. Detailed fertilization rates and application dates are provided in Table 2. Sugar beet following the rye treatment received an additional 100 kg N ha−1 due to a technical mistake. After harvest, sugar beet leaves were incorporated by 12–15 cm deep cultivator tillage before sowing winter wheat (Triticum aestivum var. “Nordkap”) in October. Trials ended with the first N fertilization of winter wheat in the following March. Crop phases for the CCs and sugar beet were defined from sowing to harvest, except for winter wheat, for which the crop phase ended at the first fertilization date.
Table 2Management dates and N fertilizer rates for CC and main crop management across the experimental trials.
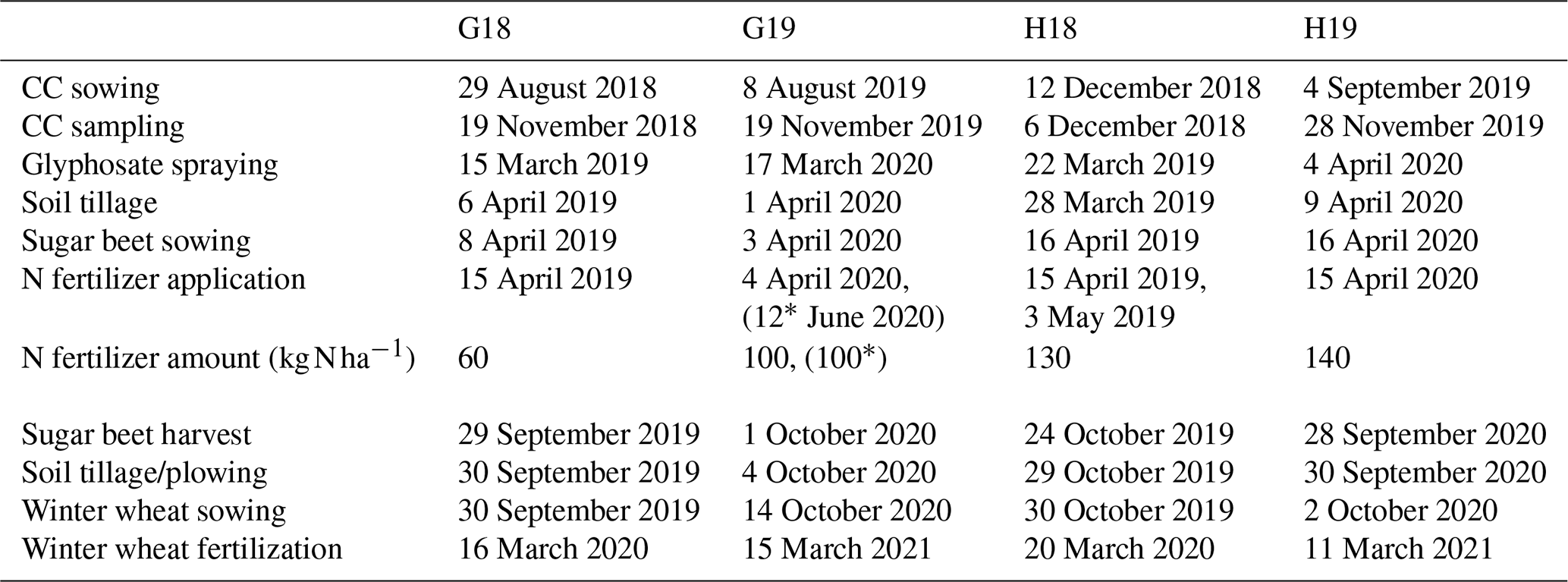
* Rye treatment only.
2.2 Plant and soil sampling, analyses, and calculations
The aboveground biomass of CCs was assessed by the end of November in each site year using four sampling points within each plot, each covering an area of 0.5 m2. To determine the dry matter (DM) content of the CC biomass, a subsample was mashed and then dried at 60 °C for 48 h. The resulting dry weight was then used to calculate the DM biomass. Elemental analysis was performed on the plant material to determine C and N concentrations, which were subsequently utilized to calculate the C and N contents of the aboveground CC biomass. Composite soil samples of five subsamples were taken biweekly from the topsoil (0–30 cm depth) using a 30 mm diameter auger. Samples were stored at −20 °C until analysis. To determine SMN content (i.e., the sum of and ), soil samples were extracted with 0.0125 M CaCl2 solution in the ratio 1:4 () and analyzed according to VDLUFA A 6.1.4.1 (VDLUFA, 2002). Gravimetric water content was measured on subsamples dried at 105 °C for 24 h. To determine soil bulk density, six undisturbed soil cores (250 cm3) per plot were taken in winter from the topsoil (3–8, 13–18, and 23–28 cm) and dried at 105 °C for 24 h before weighing. Soil bulk density was calculated according to Eq. (1):
where ρb is the bulk density (g cm−3), Ms is dry soil weight (g), and Vt is soil volume (cm3).
Water-filled pore space was calculated from the gravimetric water content and the bulk density according to Eq. (2):
where WFPS is the water-filled pore space (%), θg is the gravimetric water content (g g−1), ρb is the bulk density (g cm−3), and ρs is the assumed particle density of 2.65 g cm−3 (Hillel, 2003).
2.3 Gas sampling and analysis
The closed-chamber method was used for the N2O flux measurements, with one chamber being placed per plot. Gas samples were taken weekly or more frequently after fertilization, high precipitation, or frost-thaw events. Two types of chambers were used at the study site in Göttingen: round chambers (60 cm diameter, 45 cm height) for the CC and winter wheat phases and rectangular chambers (72 cm length, 27 cm width, 18 cm height) for the sugar beet phase. Chambers were made of white opaque PVC and sealed with rubber straps or brackets. At the study site in Hohenheim, gas fluxes were measured using chambers with an inner diameter of 30 cm as described in detail by Flessa et al. (1995). Four gas samples were taken from each chamber through a septum at 20 min intervals. Samples were collected using a 30 mL syringe and stored in pre-evacuated 12 mL vials (Exetainer, Labco Limited, UK). At both study sites, laboratory analysis employed a SCION 456-GC gas chromatograph with an electron capture detector (ECD) for N2O and a thermal conductivity detector (TCD) for CO2. Samples were introduced using a Gilson autosampler (Gilson Inc., Middleton, WI, USA). Data processing was performed using CompassCDS software. The analytical precision of the gas chromatograph was determined monthly by repeated measurements using certified standard gases (307, 760, and 6110 ppb for N2O and 201, 550, and 2500 ppm for CO2).The coefficient of variation was consistently<2 %. Mass concentrations were calculated from molar concentrations using the ideal gas equation considering the chamber temperature.
Flux rates were calculated using the gasfluxes R package (Fuss and Hueppi, 2020), selecting models based on the Akaike information criterion and kappa value. Cumulative N2O emissions for the different cropping phases were estimated using the aggfluxes function with linear interpolation between measurement dates and summed to result in the cumulative fluxes of the entire trials. Potentially mitigated indirect N2O emissions were estimated by multiplying the late autumn N uptake of CC shoots by the IPCC (2019) factors: the N2O emission factor for indirect emissions due to N leaching and runoff ( per kg N) and the factor for N losses by leaching and runoff in wet climates (FracLEACH=0.24 kg N per kg N). CO2 equivalents (CO2-eq) were calculated by using the N2O global warming potential of 273 (IPCC, 2022).
2.4 Data and statistical analyses
Data processing and analyses were carried out in R version 4.2.2 (R Core Team, 2023). For all statistical analyses, the significance level was set to p<0.05. N2O fluxes and cumulative emissions were log 10-transformed to ensure normal distribution and variance homogeneity of the model residual. Variance homogeneity and approximate normality of residuals were assessed using diagnostic plots. Flux rates that were strongly negative (i.e., lower than ) as well as those with standard errors larger than 120 , indicating high uncertainty, were excluded. In addressing the discrepancy in fertilizer application in the G19 trial, data from rye treatment, which had received the double fertilizer dose, were included in analyses, plots, and tables specific to individual site-year evaluations. However, for comprehensive analyses that combined data across all site years, the rye treatment data from G19 during the sugar beet and winter wheat phases were excluded to maintain consistency and comparability across the study. ANOVA was performed on linear models for treatment differences, with Tukey HSD tests for post hoc comparisons. A generalized least-squares regression model assessed CC impact on cumulative N2O emissions, as well as average SMN and WFPS across site years, using the nlme package (Pinheiro et al., 2023). Variance heterogeneity was addressed using the variance structure (Zuur et al., 2009), applied when found to be significant. Upon identifying a significant CC treatment effect via ANOVA, pairwise mean comparisons, with Tukey-adjusted p values, were performed on the estimated marginal means through the emmeans package (Lenth et al., 2023). The impact of environmental variables on N2O flux was analyzed using linear mixed-effects models with the lmer function from the lme4 package (Bates et al., 2015). To ensure comparability and address differences in scale, predictors were standardized using Z-score normalization from the “scales” package (Wickham et al., 2023). Biweekly SMN and WFPS values were linearly interpolated to match the weekly N2O flux measurements. The models included standardized SMN, WFPS, soil temperature, and CC treatments as fixed effects, while the site year was incorporated as a random effect to account for variability across different sites and years.
2.5 Modeling changes in SOC stocks
The effect of CCs on SOC sequestration in a 30 cm topsoil horizon was simulated over a period of 50 years for two different crop rotations, common in German agricultural practice. Crop rotation CR1, with a CC embedded every second year, had the sequence CC/bare fallow – sugar beet – winter wheat – CC/bare fallow – silage maize – winter wheat with an application of 30 m3 digestate from biogas plants before seeding of maize. Crop rotation CR2 had the crop sequence CC/bare fallow – sugar beet – winter wheat – winter rape – winter wheat, with a CC in every fourth year. No organic fertilizer was applied in crop rotation CR2. For both crop rotations, CR1 and CR2, a control without any CC was defined in contrast to three CC scenarios with saia oat, spring vetch, and winter rye as the CC. In the control and CC scenarios, it was assumed that the aboveground crop residues for winter wheat, sugar beet, and winter oilseed rape remained in the field. Effects of CCs were quantified by subtracting modeled C stocks of the control (bare fallow instead of CC) from modeled C stocks of CC treatments. Because the models RothC and C-Tool describe soil C decomposition by first-order kinetics, modeled SOC change between scenarios is linearly dependent on differences in initial C stocks and C inputs between control and CC scenarios, meaning that in our setup frequency and biomass production of grown CCs mainly control modeled effects on SOC stocks.
The long-term potential for changes in SOC content due to the cultivation of CCs was estimated using a model ensemble (Seitz et al., 2023) consisting of the RothC (Coleman and Jenkinson, 1996) and C-Tool (Taghizadeh-Toosi et al., 2014) models implemented in R using the SoilR package (Sierra et al., 2012) in combination with three allometric functions for calculating the C input from shoot and root residues of main crops as a function of yields (Franko et al., 2011; Jacobs et al., 2020; Rösemann et al., 2021). If no yield information was available for main crops, documented yields from German agricultural soil inventory sites (Jacobs et al., 2020) within a radius of 50 km were used and averaged. Yields of main crops after the CC (CC scenarios) or after the alternative bare fallow period (control) were based on observed yields from experimental treatments where the main crops followed the same CCs (saia oat, spring vetch, or winter rye) or bare fallow treatment. The C inputs of the CCs were taken from measured aboveground biomass (experimental data) and root–shoot ratios, which in turn were derived based on measurements of aboveground and belowground biomass of the CCs grown at the Göttingen site. In order to derive the C input for root biomass from CCs for the horizon of interest in the 0–30 cm profile depth, an approach according to Gale and Grigal (1987) was used to describe the root distribution as a function of depth:
Here, Y is the root fraction increasing with depth, β is a parameter, and d is the depth of the profile in centimeters. Jackson et al. (1996) set β=0.961 to describe the depth distribution of roots of cropland crops (for CCs, plant-specific values of β were determined via calibration on the basis of plant-specific available data on belowground biomass at the Göttingen site; Fig. S1 and Table S6 in the Supplement). The estimated proportion of roots in the 0–30 cm depth profile was higher for winter rye (89 %) than for saia oat and spring vetch (79 % each). In relation to the profile depth of 0–30 cm considered here, this resulted in root-to-shoot ratios of 0.24, 0.13, and 0.33 for saia oat, spring vetch, and winter rye, respectively. It was assumed that C input from root exudates corresponded to 31 % of the input from root C according to Jacobs et al. (2020).
Weather data with a monthly resolution for precipitation, temperature, and global radiation (2018–2021) were obtained from DWD grid data, as they align with the experimental crop growth periods at both sites. The suitability of these data has been validated in model evaluation studies for SOC models at German permanent observation sites (Riggers et al., 2019). To match the 50-year simulation period, these time series were repeated. Model initialization was performed separately for each model in the ensemble. For RothC, pool distribution at equilibrium was determined using an analytical solution (Dechow et al., 2019), while for C-Tool, the initial pool fractions followed the approach proposed by Taghizadeh-Toosi et al. (2014).
3.1 Weather conditions and effect of CCs on soil WFPS
The experimental trials experienced marginally warmer and drier conditions in the first establishment year compared to long-term averages (Table 3). In 2018, the rainfall totals in Göttingen and Hohenheim were 430 and 526 mm, respectively. The temperature ranges were , with occasional extremes, such as a cold spell in mid-February 2021. Soil temperatures generally fluctuated between 0 and 25 °C, although several frost events were recorded at a 5 cm depth (Fig. 1). Water-filled pore space exhibited a consistent seasonal pattern across all site years, starting low in autumn, increasing through winter to peak around February, and declining in spring and summer, with occasional short-term spikes following heavy summer rainfall (Fig. 1 and Table 3).
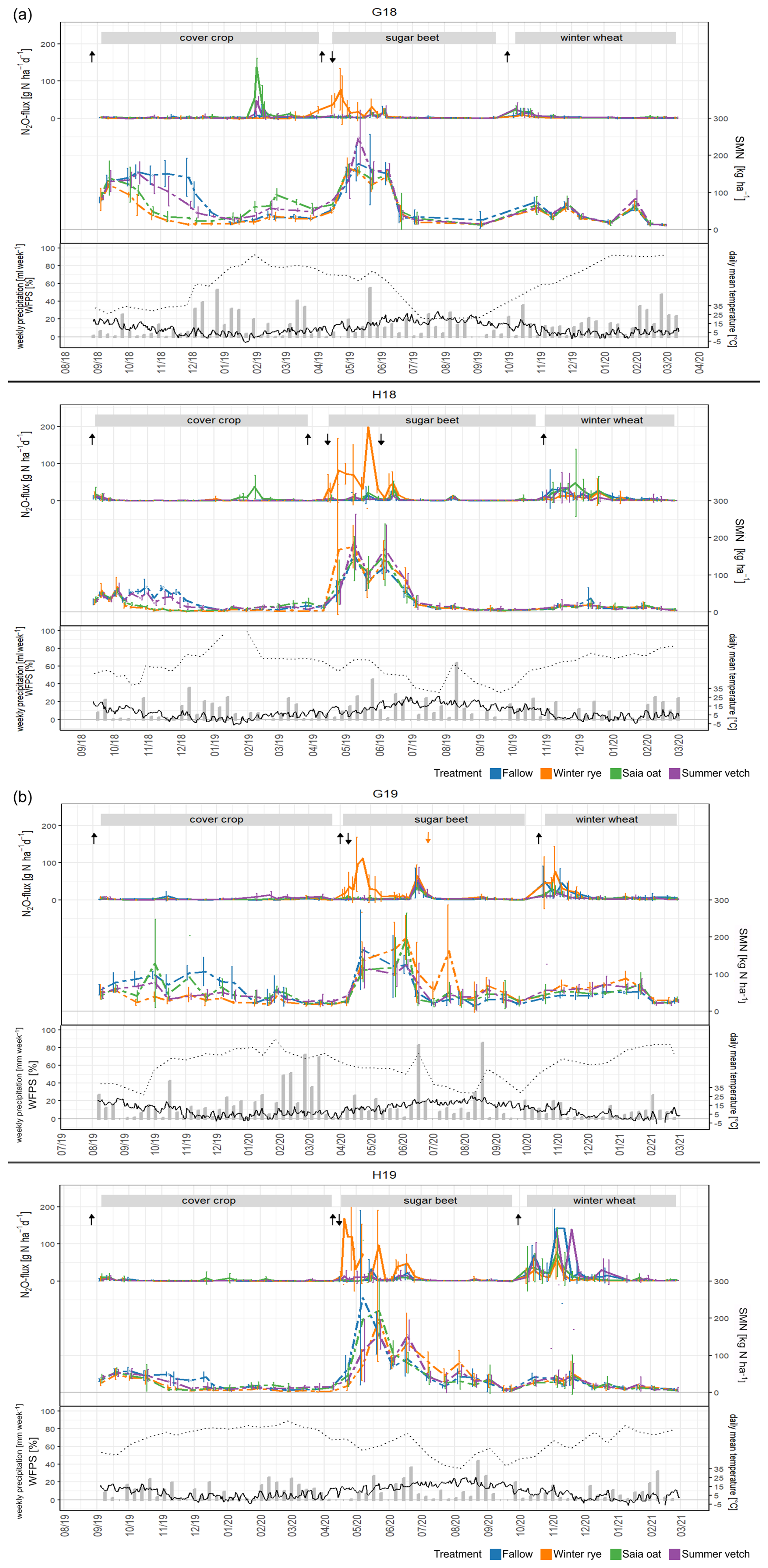
Figure 1Dynamics of N2O flux rates and soil mineral nitrogen (SMN) contents, presented as mean ± SD (n=4). Seasonal changes in topsoil water-filled pore space (WFPS%, dotted black line), daily mean air temperature (continuous black line), and weekly precipitation (gray bars) for Göttingen (upper plot) and Hohenheim (lower plot) throughout the different cropping phases for various CC treatments in (a) 2018 trials and (b) 2019 trials. Upward arrows mark soil cultivation events, while downward arrows signify N fertilization of sugar beets.
Table 3Weather conditions and soil water-filled pore space (WFPS) across different cropping phases and site years. Means ± SD. Different lowercase letters indicate statistically significant differences between site years within each column (p<0.05).
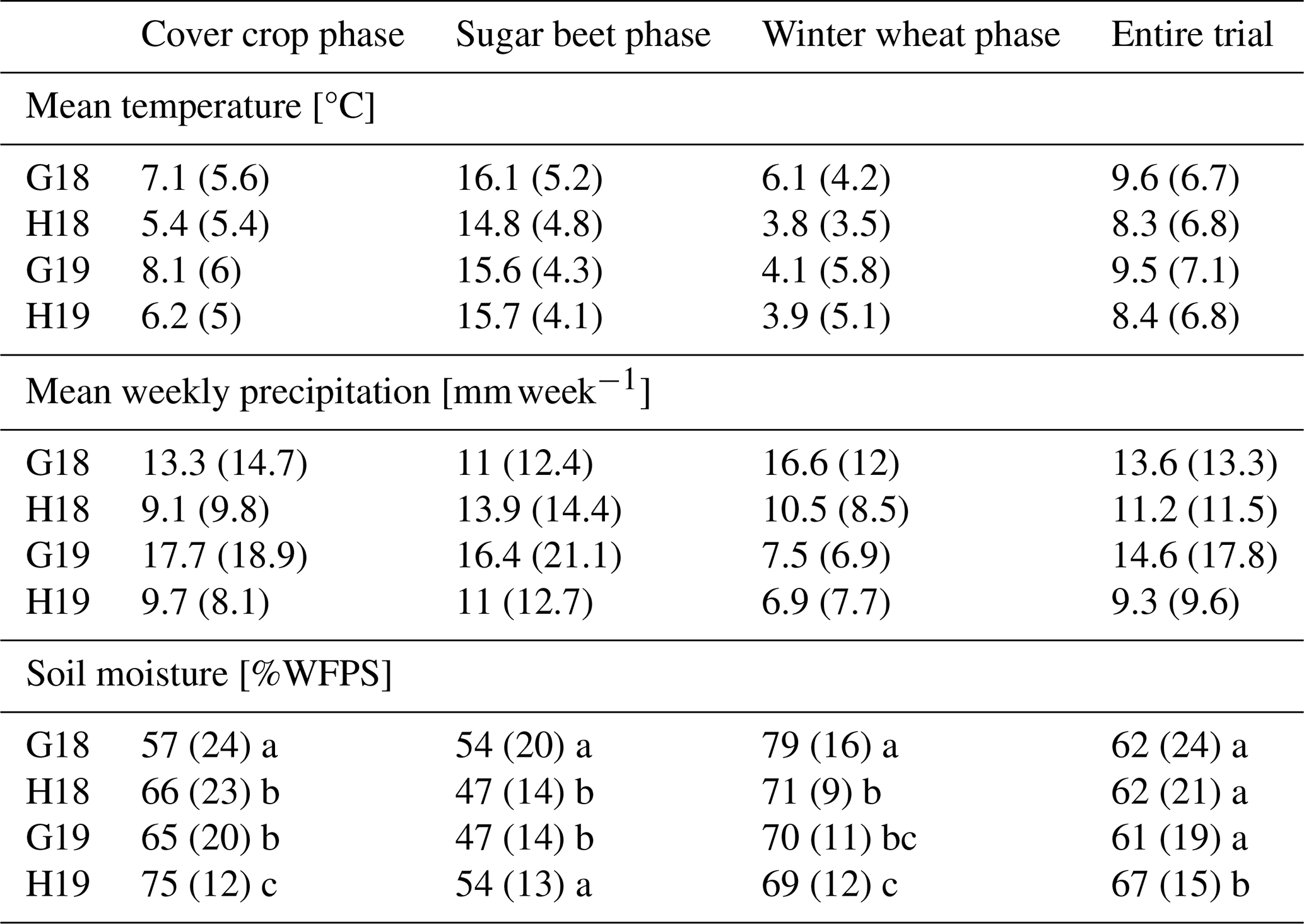
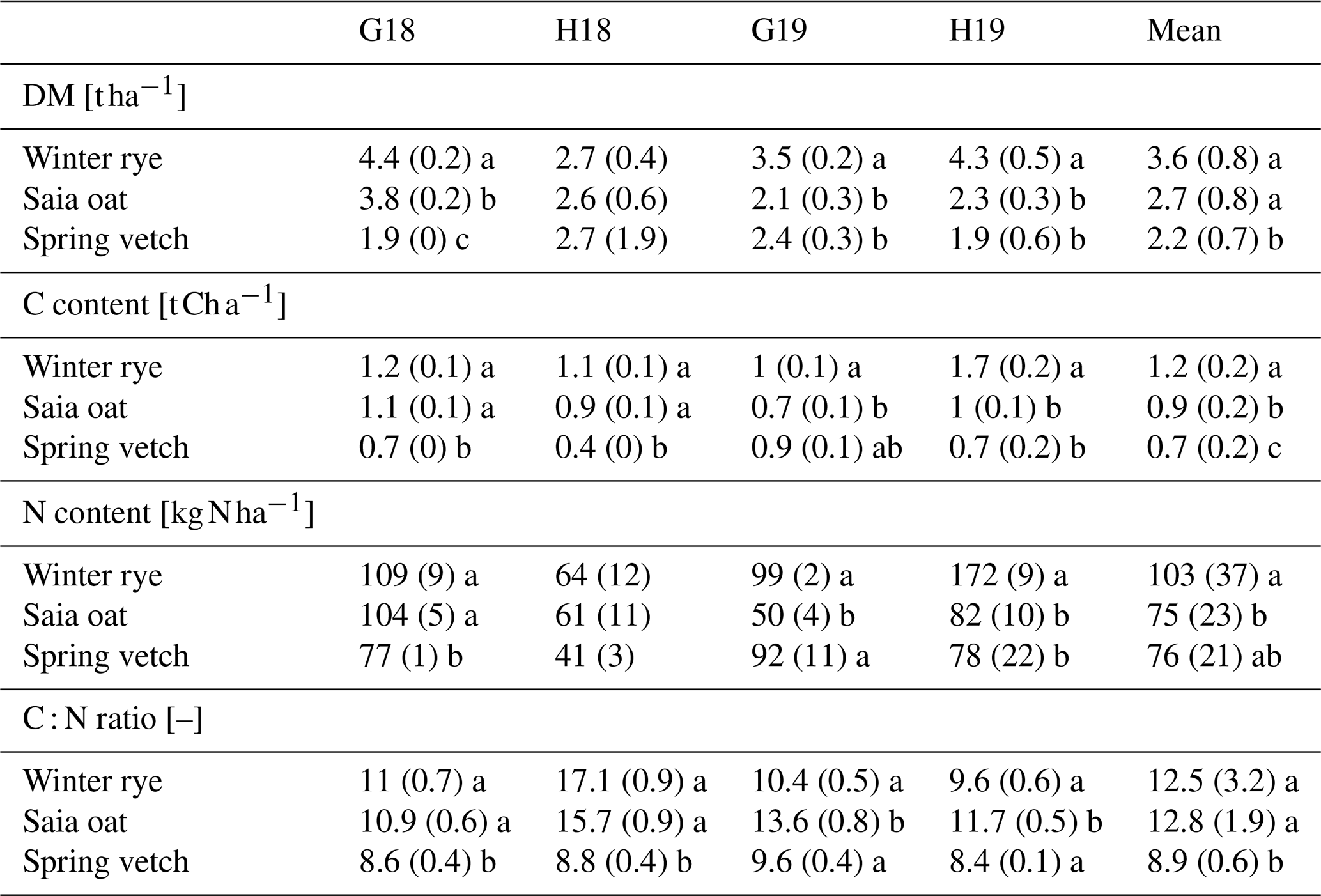
Table 4Dry matter biomass (DM), carbon (C) and nitrogen (N) contents, and C : N ratio of winter CCs in late autumn across site years. Means (n=4 for individual trials, n=16 for averages across all site years) ± SD. Significant differences between CC species within each category are denoted by different lowercase letters (p<0.05).
Concerns have been raised about the potential for increased evapotranspiration from CCs to adversely impact soil moisture (Unger and Vigil, 1998). In the G18 trial, WFPS under CC treatments was marginally lower than in fallow plots during the cover-cropping phase, though these differences were not statistically significant (Table S1 in the Supplement). The lower soil moisture observed in CC plots in G18, likely reflecting the reduced rainfall of 430 mm, supports concerns that increased evapotranspiration from CCs in semi-arid conditions (annual rainfall typically 250–500 mm) or in drier-than-average temperate years may reduce soil moisture and potentially affect subsequent crop yield (Blanco-Canqui et al., 2015; Mitchell et al., 2015). However, studies (Wang et al., 2021; Qi et al., 2011) indicate that the impact of CCs on pre-sowing soil moisture, water storage potential, and yield is highly site-specific, and any negative effects on topsoil moisture may be offset by enhanced water-holding capacity resulting from increased soil organic matter (Poeplau and Don, 2015). Indeed, long-term use of CCs such as rye has been shown to significantly improve field capacity and plant-available water (Basche et al., 2016).
3.2 Influence of CC species on SMN dynamics
Soil mineral N was predominantly composed of , with ammonium () levels remaining below 10 kg N ha−1 on most sampling dates. At the onset of trials, SMN levels varied among sites and years, with the highest average in G18 (), followed by G19 (), and the lowest in Hohenheim (29±8 and for H18 and H19, respectively). No significant differences in initial SMN levels were found between treatments within each trial (Fig. 1 and Table S2 in the Supplement).
In the present study, the cultivation of field pea as the preceding main crop led to elevated SMN levels in late summer, which were substantially higher in Göttingen than in Hohenheim. Subsequently, we observed an increase in SMN across all treatments and site years for a few weeks following the incorporation of pea straw (see Fig. 1). This trend can be attributed to the net mineralization of the pea residues, a common characteristic of legumes with low C : N ratios that promotes swift decomposition and N release (Doran and Smith, 1991).
By late autumn, significant differences in aboveground biomass DM were observed across all site years (except H18), with rye consistently achieving higher DM (3.6 t ha−1) than oat and vetch (2.7 and 2.2 t ha−1, respectively; Table 4). Correspondingly, rye exhibited the highest aboveground C content (1.2 t ha−1) and N uptake (103 kg N ha−1), while oat and vetch showed lower values (0.9 and 0.7 t ha−1 for C content, and 75 and 76 kg N ha−1 for N uptake, respectively). The C : N ratio of CC shoot biomass was consistently lower for vetch (<10) compared to rye and oat (Table 4). Simultaneously, SMN levels in CC treatments gradually declined during autumn while remaining elevated in fallow plots, with the decline being most pronounced for rye. By the end of the CC growing period in late November, all CC treatments exhibited significantly lower SMN than fallow across all site years (Fig. 1 and Table S2). In G18, the SMN difference between rye and fallow was approximately 130 kg N ha−1, 80 kg N ha−1 in G19, and 30 kg N ha−1 in both H18 and H19. Among the evaluated CCs, rye was most effective in reducing SMN – likely due to its higher biomass production, robust N uptake, and frost resilience. This observation aligns with finings of Thapa et al. (2018), who reported a positive correlation between CC biomass and uptake. In Göttingen, the more pronounced SMN differences likely reflect the higher initial SMN compared to Hohenheim. In H18, Koch et al. (2022) found that the SMN difference across the 0–90 cm soil profile closely matched the N content in rye shoots, whereas in H19, rye shoot N exceeded the SMN difference, underscoring rye's efficient N uptake and its role in mitigating leaching. Conversely, in Göttingen (G18 and G19), rye shoot N accounted for only about half of the SMN difference, suggesting contributions from unquantified factors such as root biomass, exudates, and N immobilization. Additionally, vetch was the least effective in reducing SMN, likely due to its shallower root system (Grunwald et al., 2022) and its capacity for biological N fixation (Ramirez-Garcia et al., 2015), with winter legume CCs generally producing less biomass than summer legumes and thus exerting a diminished impact on soil N uptake (Pan et al., 2022).
From November through January, SMN levels declined consistently across all treatments, reaching their lowest values in January ( in Göttingen and in Hohenheim; Fig. 1). This decline was most pronounced in fallow plots, which began with the highest SMN levels in November. While part of the winter decrease in SMN may result from leaching beyond the topsoil, the present study did not assess the entire root zone. Nonetheless, previous research has shown that CC cultivation can reduce leaching by approximately 68 % compared to fallow (Lapierre et al., 2022; Nouri et al., 2022), highlighting the significant role of CCs in enhancing N retention.
Following the frost event in late January 2019, a notable increase in SMN was observed in the frost-sensitive oat and vetch treatments in G18, as well as in oat in H18 (Fig. 1a). By the end of the cover-cropping phase in April, plots with rye exhibited the lowest SMN levels (Fig. 1 and Table S2), although the differences were not statistically significant in most site years.
Across all site years, average SMN values during the cover-cropping phase were lowest for non-legume CCs (rye and oat) and highest for bare fallow (Fig. 2a), with rye exhibiting significantly lower SMN than fallow in every trial (Table S3 in the Supplement). This confirms the first part of hypothesis (i), demonstrating that during the cover-cropping phase, non-legume CCs reduce SMN by assimilating excess N in autumn. In contrast, vetch did not consistently reduce SMN – a finding that aligns with previous research suggesting that legumes, through their capacity for atmospheric N fixation, rely less on soil N uptake (Daryanto et al., 2018; Helfrich et al., 2024; Ramirez-Garcia et al., 2015).
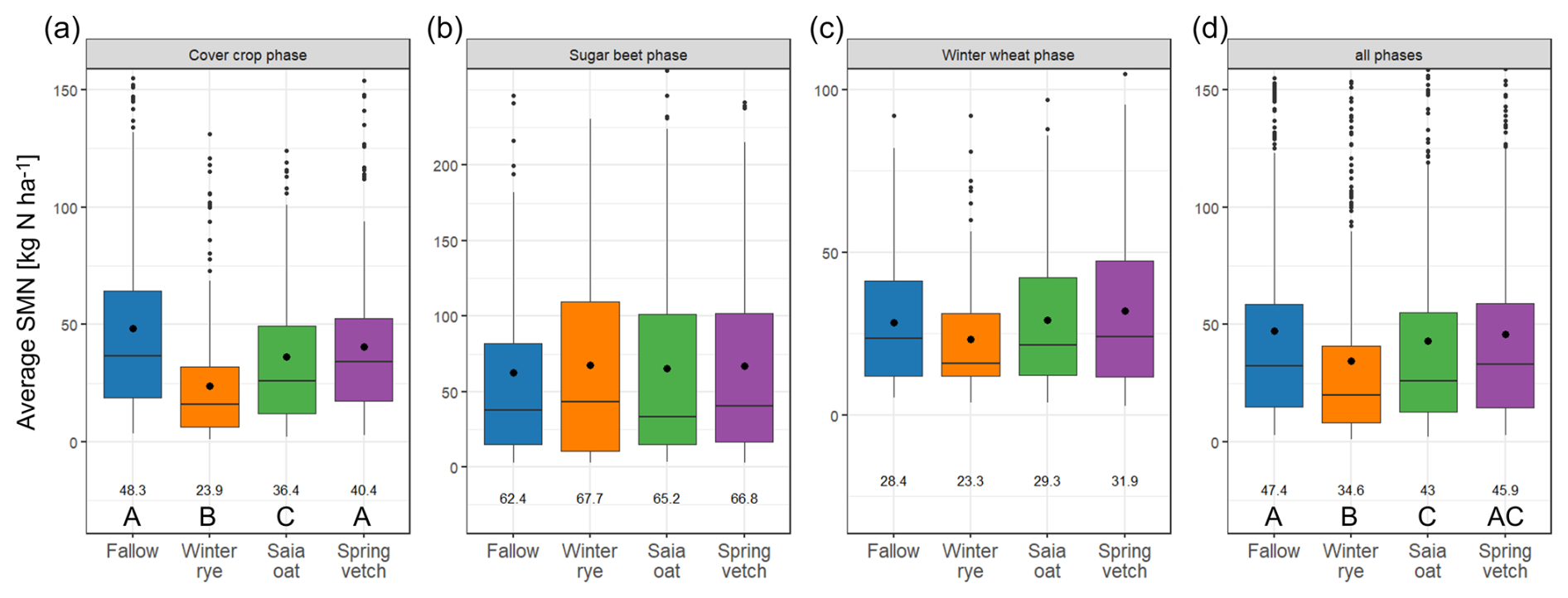
Figure 2Mean soil mineral nitrogen (SMN) during the different cropping phases (n=12 for rye treatment in the sugar beet, winter wheat, and all phases together; n=16 for all other treatments). Horizontal lines represent the median, and large dots and numbers at the bottom show the mean. Uppercase letters indicate significant differences in Z-standardized SMN values between the treatments (p<0.05).
Subsequent to the incorporation of CC residue, sugar beet sowing, and N fertilization in April, SMN levels increased to 111–247 kg N ha−1 at the onset of the sugar beet phase, effectively masking differences among CC treatments. As sugar beet N uptake increased, SMN levels declined strongly between June and July, reaching their lowest values around September (Fig. 1 and Table S2). Subsequent N fertilization – in G19 for the rye treatment and across all treatments in H18 – elevated SMN levels, yet no significant differences among CC treatments were observed during the sugar beet phase (Fig. 2), suggesting that within a fertilized framework the effect of winter CCs on SMN becomes negligible. Subsequent to the sugar beet harvest, soil cultivation, leaf residue incorporation, and winter wheat sowing, a notable SMN increase was observed across all site years, with higher levels recorded in Göttingen compared to Hohenheim (Fig. 1 and Table S3). Over the course of the entire trial, average SMN levels were highest in G18, intermediate in G19, and lowest in both H18 and H19 (Table S3). Non-legume CC treatments resulted in significantly lower SMN than bare fallow, with the rye treatment exhibiting the lowest average SMN (Fig. 2 and Table S3). These findings underscore the notion that N fertilization can supersede the impact of winter CCs on SMN. However, in settings devoid of fertilization, CCs may exert a more pronounced influence on regulating soil N availability and yield (Koch et al., 2022; Kühling et al., 2023).
3.3 Influence of CC species on N2O flux rates and cumulative N2O emissions
Across all site years, spatial and temporal variations in N2O flux rates were observed, with heavy rainfall and elevated WFPS frequently triggering emission peaks (Fig. 1). The availability of SMN in the forms of and (which serve as substrates for nitrification and denitrification) and appropriate soil moisture and temperatures that promote microbial activity are crucial factors influencing N2O production in agricultural soils (Signor and Cerri, 2013). Although the initial weeks of the trials exhibited elevated SMN levels and adequately warm soil temperatures, high N2O flux rates were not observed, a discrepancy likely due to the low WFPS recorded during late summer (Signor and Cerri, 2013). This observation is consistent with the findings of Cosentino et al. (2013), who posited that WFPS values below 59 % impede N2O production. Smith et al. (2003) observed an increase in N2O flux rates with rising WFPS, suggesting that when SMN is not a limiting factor, N2O production is enhanced. Throughout the cover-cropping phase, N2O flux rates remained relatively low, a phenomenon attributed to suboptimal microbial conditions during autumn and winter – specifically reduced temperatures and limited availability of C and N. Cosentino et al. (2013) emphasized that soil temperatures below 14 °C suppress N2O emissions, and Rummel et al. (2021) demonstrated that increased soil moisture does not boost N2O emissions when is limited. An added benefit of cultivating frost-sensitive CCs is their natural termination under appropriate winter conditions, which eliminates the need for chemical termination via herbicides. However, the termination stage significantly influences the mineralization rate of plant residues and their potential to promote N2O emissions, with immature residues leading to higher emissions (Abalos et al., 2022). In the present study, following a frost event in late January 2019, frost-sensitive oat treatments in G18 and H18 exhibited pronounced N2O emission peaks, reaching 137 (±24) in G18, with a smaller yet significant peak observed in the vetch treatment in G18. The more severe and prolonged frost in G18 resulted in greater damage to CCs in Göttingen, subsequently leading to higher N2O emission peaks. These findings are consistent with those reported by Wagner-Riddle et al. (2017), who found that cumulative annual N2O emissions in cold regions are closely linked to the number of freezing-degree days. This highlights the critical impact of temperature dynamics on N2O emissions.
The increase in N2O emissions from frost-sensitive CCs during winter can be attributed to the degradation of frost-damaged organic matter. This organic matter undergoes accelerated microbial decomposition, increases N availability, and promotes microbial respiration. These processes may create anoxic microsites favorable for N2O production (Beauchamp et al., 1989; Chen et al., 2013; Kravchenko et al., 2017). However, during milder winters, such as observed in the H19 trial, CCs persisted through the season, leading to no notable differences in cumulative N2O emissions among the treatments during the cover-cropping phase (Table S4 in the Supplement). This suggests that without early frost termination, frost-sensitive CCs share comparable effects on N2O emissions with their frost-resistant counterparts. This observation aligns with the findings of Storr et al. (2021), who determined that frost-sensitive CC species may not always terminate under temperate climates, but they continue to provide a steady supply of available C and N as the plants senesce.
During the cover-cropping phase, the frost-sensitive oat treatment resulted in higher cumulative N2O emissions compared to other treatments in 3 out of 4 site years, with this difference reaching statistical significance in both 2018 trials (G18 and H18). Vetch recorded the highest cumulative emissions in G19 (Table S4). When data were combined across all site years, treatments involving frost-sensitive CCs – particularly oat – exhibited significantly higher cumulative N2O emissions compared to bare fallow (Fig. 3). These results confirm hypothesis (ii), indicating that frost-induced damage in sensitive CCs leads to increased N2O emissions. Despite non-legume CCs reducing SMN levels relative to fallow during the cover-cropping phase, their cumulative N2O emissions did not decrease accordingly. This finding suggests that SMN concentration alone does not govern N2O emissions; instead, a combination of factors, including temperature, water-filled pore space, and the availability of C and N, plays a critical role. Consequently, this outcome does not fully support hypothesis (i), which posited that non-legume CCs reduce SMN – and subsequent N2O emissions – relative to fallow by assimilating excess N in autumn.
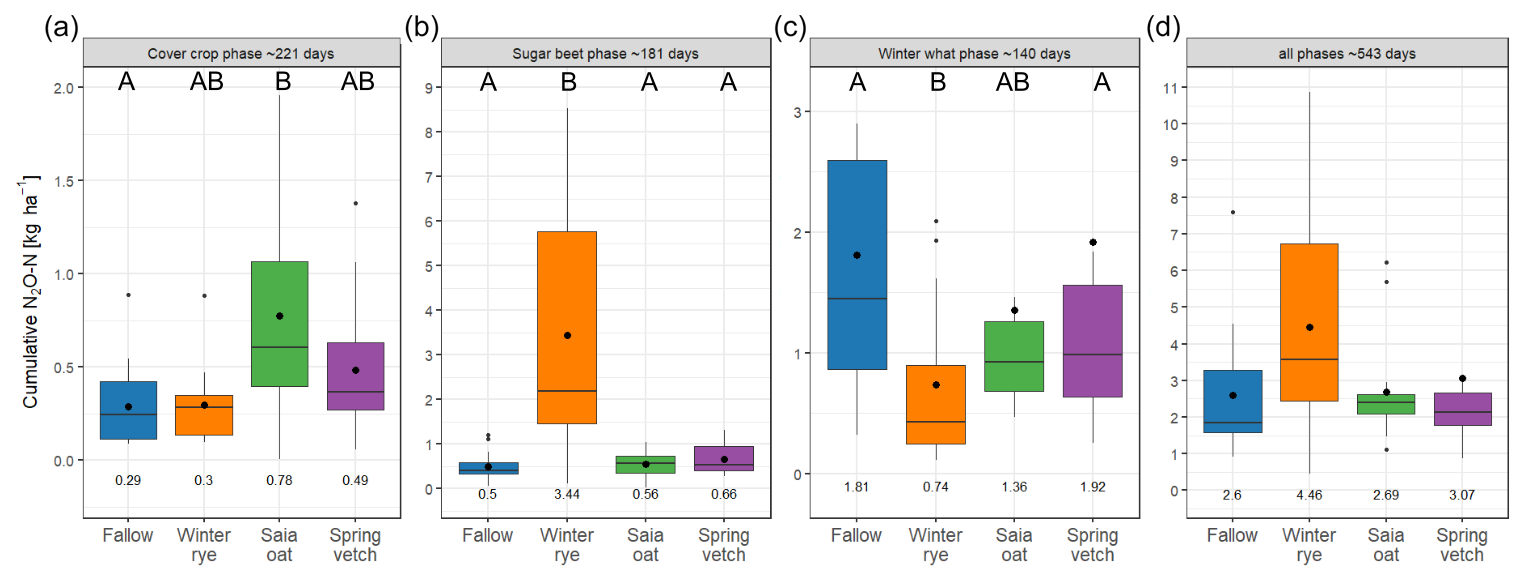
Figure 3Cumulative N2O emissions across the different cropping phases. Mean ± SD (n=12 for rye in sugar beet and winter wheat phases and all phases combined; n=16 for all other treatments). Horizontal lines represent the median, and large dots and numbers at the bottom show the mean. Uppercase letters indicate significant differences of Z-standardized cumulative N2O emission values between the treatments (p<0.05).
In our comparison of cumulative N2O emissions from frost-sensitive CCs, we found that our initial hypothesis (iii) – that residues with lower C : N ratios cause higher N2O emissions – was not supported. Instead, the C and N contents, which correlated with the DM of the residues, emerged as a more reliable indicator in this study. Despite oat having significantly higher C : N ratios than vetch, it still induced higher cumulative N2O emissions when its C and N contents were higher (Table 4 and Table S4). Moreover, although differences in C : N ratios existed between oat and vetch, both fell within a range known to facilitate net mineralization and increase soil content, thereby promoting N2O losses (Li et al., 2013). This observation aligns with the findings of Millar and Baggs (2004) and Li et al. (2013), indicating that a greater release of readily available C and N from residues with similar C : N ratios results in increased microbial activity and consequently higher N2O emissions.
A substantial body of research has documented a rapid increase in N2O emissions following N fertilizer application, primarily due to enhanced substrate availability for nitrification and denitrification (Dobbie and Smith, 2003; Weitz et al., 2001). Similarly, the addition of organic amendments to improve soil fertility and crop productivity can stimulate N2O emissions through mechanisms such as the priming effect (Thangarajan et al., 2013). The incorporation of CC residues has also been linked to significant increases in N2O emissions (Abalos et al., 2022), with Mutegi et al. (2010) attributing 60 % of annual N2O emissions to tillage and residue incorporation. In this study, the incorporation of CC residues combined with sugar beet sowing and N fertilization resulted in elevated N2O flux rates lasting 6 to 8 weeks across all treatments. Interestingly, despite comparable SMN levels across treatments due to N fertilization, the rye treatment exhibited significantly higher cumulative N2O emissions during the sugar beet phase (3.4 ) compared to fallow, oat, and vetch (approximately 0.5–0.66 ). These results support hypothesis (iv), demonstrating that incorporating CC residues with higher biomass leads to increased N2O emissions. Unlike the shallow tilling (15 cm) applied in most treatments, rye required deep plowing (up to 30 cm) to manage its extensive root system, which may have further influenced N2O emissions by disturbing soil organic biomass and enhancing mineralization rates. Observations from the Göttingen trials suggest that variations in incorporated CC biomass are the primary drivers behind these differences; the larger biomass of fresh rye residues likely resulted in increased C turnover, heightened microbial activity, more rapid oxygen depletion, and the formation of anaerobic microsites that favored N2O production (Blagodatsky et al., 2011). Additionally, WFPS exerted a notable influence on N2O flux rates during the sugar beet phase (Table S5), with increased emissions observed following heavy rainfall events. While subsequent N fertilization in the rye treatment (in G19) and across all treatments in H18 raised SMN levels, N2O flux rates were much lower later in the season when WFPS had declined. From July until the end of the sugar beet phase, N2O flux rates nearly diminished to non-detectable levels, likely reflecting reduced WFPS and minimal SMN as a result of robust N uptake by the maturing sugar beet plants. According to the IPCC emissions factor for wet climates, approximately 0.6 % of the N present in crop residues is converted into N2O (IPCC, 2019). For rye, with an average shoot biomass N content of 103 kg N ha−1, this conversion would account for only about 21 % of the observed 2.9 increase compared to fallow during the sugar beet phase. Thus, additional contributions from rye root decomposition and enhanced soil organic matter decomposition likely explain the surplus N2O emissions. Li et al. (2015) observed that residues with high C : N ratios can still lead to significant N2O emissions if the C is readily decomposable, while Abalos et al. (2022) identified N and easily degradable organic fractions as key factors affecting N2O emissions from crop residues. Therefore, the elevated cumulative N2O emissions following rye incorporation can be attributed to its high biomass yield and the large amounts of readily decomposable C and N from its residues.
After the sugar beet harvest, subsequent soil cultivation, leaf residue incorporation, and winter wheat sowing led to an increase in N2O fluxes. During the winter wheat phase, cumulative N2O emissions were significantly lower in the rye treatment than in the fallow and vetch treatments, even though no residual effect of the CCs on SMN levels was observed. Furthermore, SMN levels had a significant effect on N2O fluxes during the winter wheat phase (Table S5), indicating that SMN might have been the limiting factor for N2O production in this phase.
The majority of studies investigating CCs in agricultural rotations have focused on short-term N2O emissions during either CC growth or following residue incorporation, leaving a knowledge gap regarding year-round N2O emissions in systems incorporating CC cultivation (Muhammad et al., 2019). In the current study, even though non-legume CCs consistently produced lower SMN values than bare fallow over the entire trial period – with rye showing the lowest SMN levels – rye resulted in the highest total cumulative N2O emissions, followed by oat and vetch, while fallow exhibited the lowest emissions. Nevertheless, these differences were not statistically significant over the entire trial period. These findings are consistent with those of Basche et al. (2014), who reported that CCs tend to have a net neutral effect on N2O emissions when measured over long periods, highlighting the importance of long-term measurements to better understand the full impact of CCs on N2O dynamics.
Despite higher SMN levels in Göttingen, cumulative N2O emissions over the entire trial were generally greater in the Hohenheim trials – highest in H19 and lowest in G18 – although these differences were not statistically significant (Table S4). This discrepancy may be attributed to the higher clay content in Hohenheim soils, which likely reduced gas diffusivity, promoted the formation of anaerobic microsites, and consequently elevated denitrification rates – a factor known to increase N2O emissions (Bollmann and Conrad, 1998; Pelster et al., 2012). These findings underscore the importance of soil type and climatic conditions in regulating N2O emissions.
3.4 Mitigation potential of indirect N2O emissions by non-legume CCs
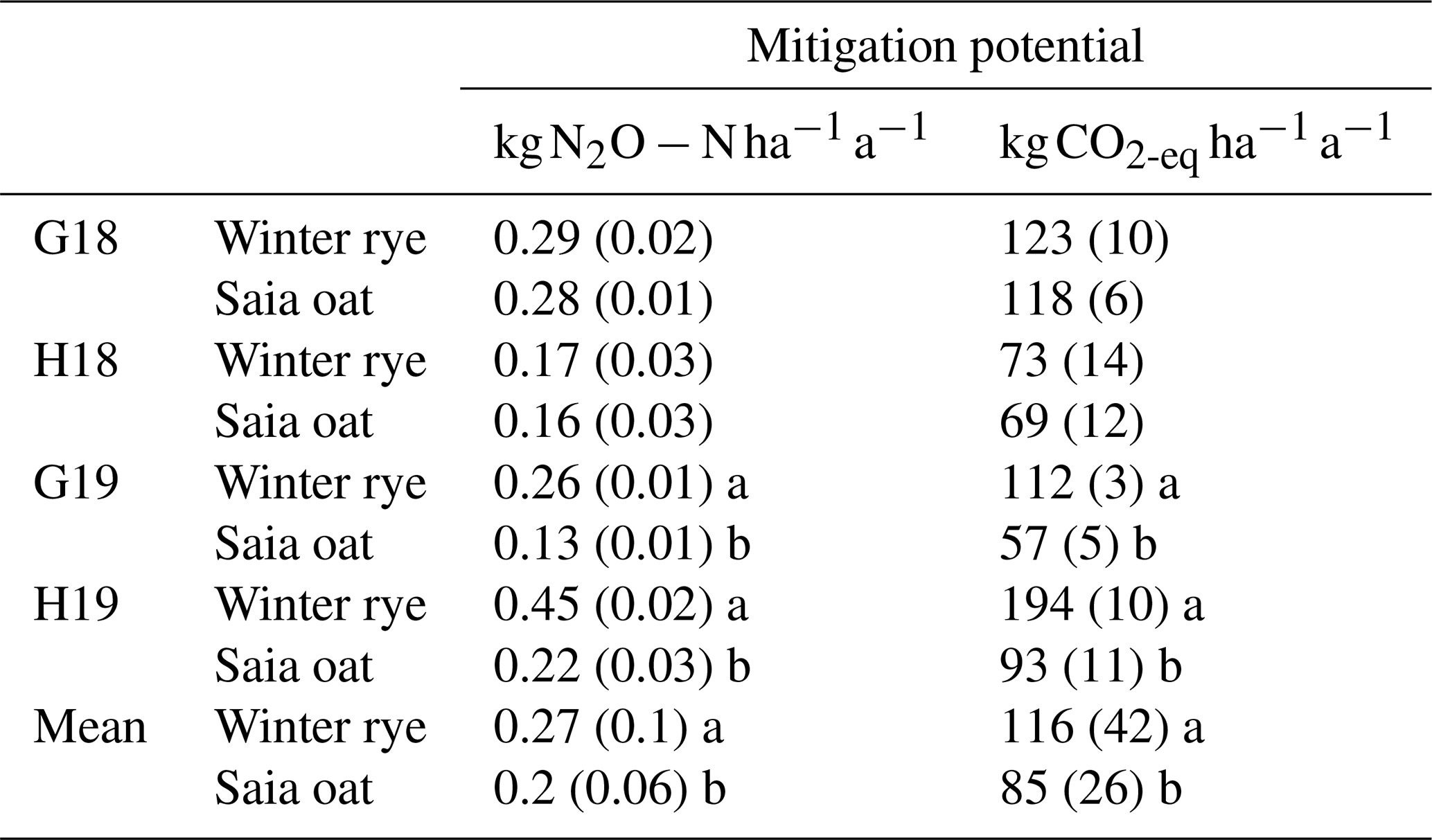
The mitigation potential for indirect N2O emissions induced by leaching – calculated from non-legume CC N uptake in late autumn – was significantly higher in rye, averaging 0.27 (approximately 116 ). Oat reduced emissions by an average of 0.2 (approximately 85 ; Table 5). It should be noted that vetch was excluded from the calculation due to uncertainties in distinguishing soil N uptake from N biologically fixed. The cultivation of CCs, particularly during periods of high soil N availability and precipitation, has been demonstrated to absorb excess N and thereby reduce leaching, mitigating indirect N2O emissions. Although CC cultivation in this study did not diminish direct N2O emissions – and, in the case of rye, slightly increased them – the observed potential reduction in leaching suggests a potential to lower indirect N2O emissions. This finding is in line with Parkin et al., 2016), who reported that rye, while neutral regarding direct N2O emissions, substantially decreased indirect N2O emissions over a decade-long trial. However, the methodology employed here, based on N content in CC aboveground biomass, provides an approximate estimate of the potential mitigation of indirect N2O emissions. It does not account for soil water movement, N concentrations in leachate, N stored in roots, or the N mineralization occurring during the winter period. Furthermore, mitigation levels likely vary with soil type and climatic factors, with sandy soils and periods of high precipitation exhibiting more pronounced effects (Simmelsgaard, 1998). Thus, these results represent an oversimplification of the complex processes involved and should be interpreted with caution.
Table 5Mitigation potential of non-legume cover crops (CCs) in reducing indirect N2O emissions induced by N leaching derived from pre-winter N uptake of CCs. Mean values (n=4 for individual trials, n=16 for overall averages) ± SD. Statistically significant differences between rye and oat within each trial and overall averages are denoted by different lowercase letters.
3.5 Effect of CCs on long-term soil C sequestration
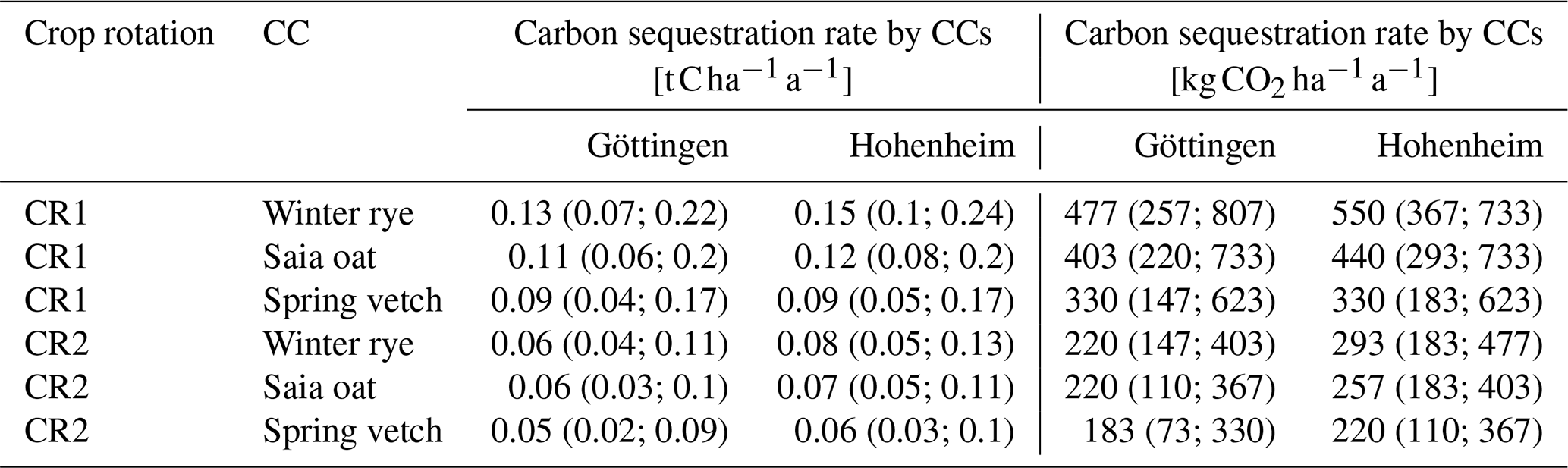
According to our model results, soil C sequestration from winter rye, saia oat, and spring vetch significantly contributes to greenhouse gas mitigation by CCs and is a relevant sink compared to direct and indirect N2O fluxes. At the Göttingen sites, average C inputs from main crops in the control scenarios (no CC) were about 4.15 and 4.51 for crop rotations CR1 and CR2, respectively, whereas at the Hohenheim site they were approximately 10 % lower (3.76 and 4.15 ; Table S7 in the Supplement). CR1 exhibited lower C inputs than CR2, primarily due to a higher share of winter crops that maintain larger amounts of incorporated residue, even though CCs were grown at a 2-year interval in CR1 compared to every 4 years in CR2. Among the CCs, winter rye exhibited the highest C inputs, providing additional increases of 20 %–24 % in CR1 % and 9 %–11 % in CR2. Saia oat followed, with increases of 15 %–17 % in CR1 % and 7 %–8 % in CR2, and spring vetch had the lowest increases, with 11 %–12 % in CR1 % and 5 % in CR2 (Table S8 in the Supplement). As anticipated, the rate of C sequestration was observed to be approximately double that of the 4-year cultivation interval (CR2) in the 2-year cultivation interval (CR1). In CR1, the annual C sequestration rates averaged for the simulation periods of 50 years were highest for winter rye (0.13 and 0.15 for Göttingen Hohenheim, respectively), followed by saia oat (0.11 and 0.12 for Göttingen and Hohenheim, respectively) and spring vetch (0.09 for both sites, Table 6 and Fig. 4). Sequestration rates found here for a 30 cm profile are of the same order as or slightly lower than those reported in several meta-studies (Abdalla et al., 2019; Blanco-Canqui, 2022; Bolinder et al., 2020; Poeplau and Don, 2015). For example, Poeplau and Don (2015) reported an averaged sequestration rate of in the 0–22 cm depth interval. Assuming proportionality between C input and soil C stock changes and a uniform SOC distribution in the plow horizon, this would translate to sequestration rates of for a cover crop grown every second year and for one grown every fourth year. These CC-related effects are higher than those obtained in our modeling study (0.09–0.15 for CR1 and 0.05–0.08 for CR2). This is consistent with lower C inputs from CCs in this study (CR1: 0.82–1.82 ) compared to reported mean C inputs from CCs of 1.87 in Poeplau and Don (2015). One likely reason for the lower C inputs in Göttingen and Hohenheim is the relatively late sowing of cover crops (end of August to early September), which hampered optimal biomass production.
Table 6Simulated carbon sequestration rates caused by cover crops (CCs) averaged over a simulation period of 50 years, with the model structural uncertainties in parentheses.
However, the study's findings are subject to certain uncertainties because the models employed are pure soil C models. First, plant growth was estimated from experimental data rather than dynamically simulated, which may limit the interpretation of the results. Second, the representation of CC biomass was based on only 2 relatively dry years, potentially underestimating the full variability in C inputs. Moreover, N availability, which may affect biomass growth and C utilization efficiency during the conversion of crop residues to soil organic matter (Jian et al., 2020), is not considered in the current models.
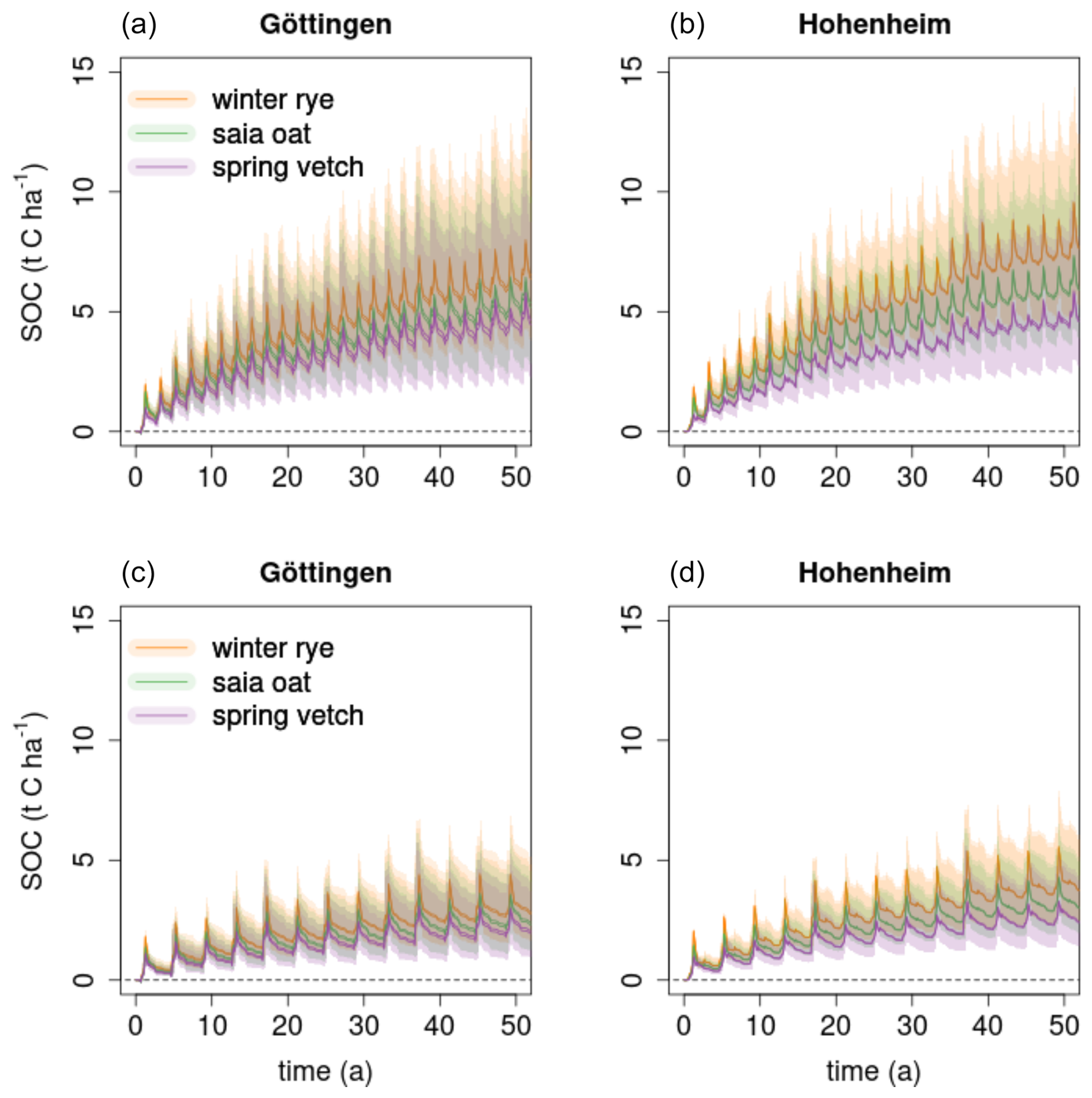
Figure 4Modeled effect of cover crops (CCs) on the increase in SOC stocks (0–30 cm) compared to the control without CC for regionally common crop rotations (a, b) CR1, which is “CC/bare fallow – sugar beet – winter wheat – CC/bare fallow – silage maize with an application of 30 m3 digestate from biogas plants before seeding of the maize”, and (c, d) CR2, which is“CC/bare fallow – sugar beet – winter wheat – winter rape – winter wheat”, with a CC in every fourth year at the Göttingen and Hohenheim experimental sites (two fields per site). Shaded areas show the variability of the model ensemble.
These considerations are crucial in selecting CCs, as they underscore the intricate relationship between N dynamics, residue decomposition, and microbial activity. This complexity highlights the need for a comprehensive understanding of CC selection and management to optimize N uptake efficiency while reducing N2O emissions. Modeling approaches could serve as valuable tools in this context by predicting the most suitable CC types based on site-specific factors such as soil type, climate, and the subsequent main crop. Moreover, conducting incubation studies under controlled conditions using labeled N can further elucidate the primary drivers of N2O emissions, thereby facilitating more informed, locally tailored CC decisions. Ultimately, achieving a balance between maximizing N capture during the cover-cropping phase and minimizing N2O emissions during residue incorporation is paramount. Additional research into the mechanisms behind elevated N2O emissions from CC residues and the development of effective mitigation strategies is essential to advance more sustainable agricultural practices.
The current study highlights the complex role of CCs in agricultural systems, particularly in relation to soil N dynamics, N2O emissions, and C sequestration. While CCs, especially non-legumes like rye and oat, have demonstrated significant potential in reducing SMN levels and mitigating the risk of leaching, their impact on N2O emissions is multifaceted. Our findings highlight that frost-sensitive CCs can lead to increased N2O emissions following frost events or, in all cases, the incorporation of crop residues for establishing the next cash crop combined with mineral N fertilization. However, the potential of CCs to mitigate indirect N2O emissions and sequester C suggests a beneficial aspect of their use in sustainable agriculture. This balance between N capture, N2O emission, and C sequestration emphasizes the need for strategic management of CCs to harness their benefits fully while minimizing potential environmental drawbacks. The type of CC (frost tolerance, legume or non-legume, and pest control aspects) in particular needs to be chosen carefully depending on the following cash crops and climatic conditions. Future research should focus on developing crop rotation and site-specific management practices that optimize CC benefits for soil health, crop productivity, and climate change mitigation.
Data are available here: https://doi.org/10.25625/HFEDA7 (Nasser, 2024).
The supplement related to this article is available online at https://doi.org/10.5194/soil-11-489-2025-supplement.
VN conceived of the research, contributed to methodology development, performed data curation, performed formal calculations as well as data analysis and investigation, and drafted the original paper. MH provided critical input during the review and editing process, contributed resources and validation, and participated in drafting the paper. RD performed formal analysis, provided visualization support, and participated in drafting the paper. AM and PSR contributed to the review and editing of the paper. HJK provided supervision throughout the project, contributed to the review and editing process, and played a role in the design of the study. RR provided feedback, supervised the project at the Hohenheim site, contributed to the conceptualization, and acquired partial funding for the research, while LE performed data collection and curation at the Hohenheim site. KD provided feedback, supervised the project at the Göttingen site, contributed to the conceptualization, and acquired partial funding for the research. All authors reviewed and approved the final version of the paper.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
This article is part of the special issue “Trade-offs and synergies of soil carbon sequestration and environmental impacts: implications for agricultural management”. It is a result of the EGU 2023, Vienna, Austria, 23–28 April 2023.
This study was conducted as part of the joint research project “Reduction of greenhouse gas emissions from crop production through site-specific optimized cover crop systems” (THG ZwiFru), funded by the German Federal Office for Agriculture and Food on behalf of the Federal Ministry of Food and Agriculture (funding reference no. 281B200716). We extend our sincere appreciation to the local farmers in Göttingen and the crew at the Institute of Sugar Beet Research in Göttingen for facilitating our experiments on their fields. Special thanks go to Thorsten Gronemann and Marlies Niebuhr for their assistance in the field and to Dennis Grunwald for his efforts in data curation.
This project has been funded by the Federal Ministry of Food and Agriculture (BMEL), following a decision by the Parliament of the Federal Republic of Germany, and is administered through the Federal Office for Agriculture and Food (BLE) under the innovation support program (funding reference no. 281B200716). This work was partially funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany's Excellence Strategy – EXC 2070 – 390732324.
This paper was edited by Mart Ros and reviewed by Chantal Hendriks and one anonymous referee.
Abalos, D., Rittl, T. F., Recous, S., Thiébeau, P., Topp, C. F. E., van Groenigen, K. J., Butterbach-Bahl, K., Thorman, R. E., Smith, K. E., Ahuja, I., Olesen, J. E., Bleken, M. A., Rees, R. M., and Hansen, S.: Predicting field N2O emissions from crop residues based on their biochemical composition: A meta-analytical approach, Sci. Total Environ., 812, 152532, https://doi.org/10.1016/j.scitotenv.2021.152532, 2022.
Abdalla, M., Hastings, A., Cheng, K., Yue, Q., Chadwick, D., Espenberg, M., Truu, J., Rees, R. M., and Smith, P.: A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity, Glob. Change Biol., 25, 2530–2543, https://doi.org/10.1111/gcb.14644, 2019.
Adetunji, A. T., Ncube, B., Mulidzi, R., and Lewu, F. B.: Management impact and benefit of cover crops on soil quality: A review, Soil Till. Res., 204, 104717, https://doi.org/10.1016/j.still.2020.104717, 2020.
Araújo, F. C. D., Nascente, A. S., Guimarães, J. L. N., Sousa, V. S., Freitas, M. A. M. D., and Santos, F. L. D. S.: Cover crops in the off-season in the weed management at notillage area, Rev. Caatinga, 34, 50–57, https://doi.org/10.1590/1983-21252021v34n106rc, 2021.
Balkcom, K. S., Duzy, L. M., Kornecki, T. S., and Price, A. J.: Timing of Cover Crop Termination: Management Considerations for the Southeast, Crop, Forage and Turfgrass Management, 1, cftm2015.0161, https://doi.org/10.2134/cftm2015.0161, 2015.
Basche, A. D., Miguez, F. E., Kaspar, T. C., and Castellano, M. J.: Do cover crops increase or decrease nitrous oxide emissions? A meta-analysis, J. Soil Water Conserv., 69, 471–482, https://doi.org/10.2489/jswc.69.6.471, 2014.
Basche, A. D., Kaspar, T. C., Archontoulis, S. V., Jaynes, D. B., Sauer, T. J., Parkin, T. B., and Miguez, F. E.: Soil water improvements with the long-term use of a winter rye cover crop, Agr. Water Manage., 172, 40–50, https://doi.org/10.1016/j.agwat.2016.04.006, 2016.
Bates, D., Mächler, M., Bolker, B., and Walker, S.: Fitting Linear Mixed-Effects Models Using lme4, J. Stat. Softw., 67, 1–48, https://doi.org/10.18637/jss.v067.i01, 2015.
Beauchamp, E. G., Trevors, J. T., and Paul, J. W.: Carbon Sources for Bacterial Denitrification, in: Advances in Soil Science, vol. 10, edited by: Stewart, B. A., Springer, New York, NY, 113–142, https://doi.org/10.1007/978-1-4613-8847-0_3, 1989.
Blagodatsky, S., Grote, R., Kiese, R., Werner, C., and Butterbach-Bahl, K.: Modelling of microbial carbon and nitrogen turnover in soil with special emphasis on N-trace gases emission, Plant Soil, 346, 297–330, https://doi.org/10.1007/s11104-011-0821-z, 2011.
Blanco-Canqui, H.: Cover crops and carbon sequestration: Lessons from U. S. studies, Soil Sci. Soc. Am. J., 86, 501–519, https://doi.org/10.1002/saj2.20378, 2022.
Blanco-Canqui, H., Shaver, T. M., Lindquist, J. L., Shapiro, C. A., Elmore, R. W., Francis, C. A., and Hergert, G. W.: Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils, Agron. J., 107, 2449–2474, https://doi.org/10.2134/agronj15.0086, 2015.
Bolinder, M. A., Crotty, F., Elsen, A., Frac, M., Kismányoky, T., Lipiec, J., Tits, M., Tóth, Z., and Kätterer, T.: The effect of crop residues, cover crops, manures and nitrogen fertilization on soil organic carbon changes in agroecosystems: a synthesis of reviews, Mitig. Adapt. Strat. Gl., 25, 929–952, https://doi.org/10.1007/s11027-020-09916-3, 2020.
Bollmann, A. and Conrad, R.: Influence of O2 availability on NO and N2O release by nitrification and denitrification in soils, Glob. Change Biol., 4, 387–396, https://doi.org/10.1046/j.1365-2486.1998.00161.x, 1998.
Chatskikh, D., Olesen, J. E., Hansen, E. M., Elsgaard, L., and Petersen, B. M.: Effects of reduced tillage on net greenhouse gas fluxes from loamy sand soil under winter crops in Denmark, Agr. Ecosyst. Environ., 128, 117–126, https://doi.org/10.1016/j.agee.2008.05.010, 2008.
Chen, H., Li, X., Hu, F., and Shi, W.: Soil nitrous oxide emissions following crop residue addition: a meta-analysis, Glob. Change Biol., 19, 2956–2964, https://doi.org/10.1111/gcb.12274, 2013.
Coleman, K. and Jenkinson, D. S.: RothC-26.3 – A Model for the turnover of carbon in soil, in: Evaluation of Soil Organic Matter Models, NATO ASI Series, vol. 38, edited by: Powlson, D. S., Smith, P., and Smith, J. U., Springer, Berlin, Heidelberg, Berlin, Heidelberg, 237–246, https://doi.org/10.1007/978-3-642-61094-3_17, 1996.
Constantin, J., Beaudoin, N., Laurent, F., Cohan, J.-P., Duyme, F., and Mary, B.: Cumulative effects of catch crops on nitrogen uptake, leaching and net mineralization, Plant Soil, 341, 137–154, https://doi.org/10.1007/s11104-010-0630-9, 2011.
Cosentino, V. R. N., Figueiro Aureggui, S. A., and Taboada, M. A.: Hierarchy of factors driving N2O emissions in non-tilled soils under different crops, Eur. J. Soil Sci., 64, 550–557, https://doi.org/10.1111/ejss.12080, 2013.
Daryanto, S., Fu, B., Wang, L., Jacinthe, P.-A., and Zhao, W.: Quantitative synthesis on the ecosystem services of cover crops, Earth-Sci. Rev., 185, 357–373, https://doi.org/10.1016/j.earscirev.2018.06.013, 2018.
Davidson, E. A.: The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860, Nat. Geosci., 2, 659–662, https://doi.org/10.1038/ngeo608, 2009.
Dechow, R., Franko, U., Kätterer, T., and Kolbe, H.: Evaluation of the RothC model as a prognostic tool for the prediction of SOC trends in response to management practices on arable land, Geoderma, 337, 463–478, https://doi.org/10.1016/j.geoderma.2018.10.001, 2019.
Dobbie, K. E. and Smith, K. A.: Nitrous oxide emission factors for agricultural soils in Great Britain: the impact of soil water-filled pore space and other controlling variables, Glob. Change Biol., 9, 204–218, https://doi.org/10.1046/j.1365-2486.2003.00563.x, 2003.
Doran, J. W. and Smith, M. S.: Role of cover crops in nitrogen cycling, Cover crops for clean water, 85, 85–90, 1991.
DWD: Wetter und Klima – Deutscher Wetterdienst – CDC (Climate Data Center), https://www.dwd.de/EN/climate_environment/cdc/cdc_node_en.html (last access: 12 July 2023), 2023.
Elhakeem, A., van der Werf, W., Ajal, J., Lucà, D., Claus, S., Vico, R. A., and Bastiaans, L.: Cover crop mixtures result in a positive net biodiversity effect irrespective of seeding configuration, Agr. Ecosyst. Environ., 285, 106627, https://doi.org/10.1016/j.agee.2019.106627, 2019.
Finney, D. M., Buyer, J. S., and Kaye, J. P.: Living cover crops have immediate impacts on soil microbial community structure and function, J. Soil Water Conserv., 72, 361–373, https://doi.org/10.2489/jswc.72.4.361, 2017.
Flessa, H., Dörsch, P., and Beese, F.: Seasonal variation of N2O and CH4 fluxes in differently managed arable soils in southern Germany, J. Geophys. Res.-Atmos., 100, 23115–23124, https://doi.org/10.1029/95JD02270, 1995.
Fosu, M., Kühne, R. F., and Vlek, P. L. G.: Mineralization and microbial biomass dynamics during decomposition of four leguminous residues, Journal of Biological Sciences, 7, 632–637, 2007.
Franko, U., Kolbe, H., Thiel, E., and Ließ, E.: Multi-site validation of a soil organic matter model for arable fields based on generally available input data, Geoderma, 166, 119–134, https://doi.org/10.1016/j.geoderma.2011.07.019, 2011.
Fuss, R. and Hueppi, R.: gasfluxes: Greenhouse Gas Flux Calculation from Chamber Measurements, CRAN [code], https://git-dmz.thuenen.de/fuss/gasfluxes (last access: 11 February 2023), 2020.
Gabriel, J. L. and Quemada, M.: Replacing bare fallow with cover crops in a maize cropping system: Yield, N uptake and fertiliser fate, Eur. J. Agron., 34, 133–143, https://doi.org/10.1016/j.eja.2010.11.006, 2011.
Gale, M. R. and Grigal, D. F.: Vertical root distributions of northern tree species in relation to successional status, Can. J. Forest. Res., 17, 829–834, https://doi.org/10.1139/x87-131, 1987.
Galloway, J. N. and Cowling, E. B.: Reactive Nitrogen and The World: 200 Years of Change, ambi, 31, 64–71, https://doi.org/10.1579/0044-7447-31.2.64, 2002.
German Fertilizer Ordinance: Verordnung zur Neuordnung der guten fachlichen Praxis beim Düngen, Bundesgesetzblatt Teil I, 1305, 2017.
Goodroad, L. L. and Keeney, D. R.: Nitrous oxide emissions from soils during thawing, Can. J. Soil. Sci., 64, 187–194, https://doi.org/10.4141/cjss84-020, 1984.
Grunwald, D., Stracke, A., and Koch, H.: Cover crop effects on soil structure and early sugar beet growth, Soil Use Manage., 39, 209–217, https://doi.org/10.1111/sum.12800, 2022.
Guenet, B., Gabrielle, B., Chenu, C., Arrouays, D., Balesdent, J., Bernoux, M., Bruni, E., Caliman, J.-P., Cardinael, R., Chen, S., Ciais, P., Desbois, D., Fouche, J., Frank, S., Henault, C., Lugato, E., Naipal, V., Nesme, T., Obersteiner, M., Pellerin, S., Powlson, D. S., Rasse, D. P., Rees, F., Soussana, J.-F., Su, Y., Tian, H., Valin, H., and Zhou, F.: Can N2O emissions offset the benefits from soil organic carbon storage?, Glob. Change Biol., 27, 237–256, https://doi.org/10.1111/gcb.15342, 2021.
Hanrahan, B. R., King, K. W., Duncan, E. W., and Shedekar, V. S.: Cover crops differentially influenced nitrogen and phosphorus loss in tile drainage and surface runoff from agricultural fields in Ohio, USA, J. Environ. Manage., 293, 112910, https://doi.org/10.1016/j.jenvman.2021.112910, 2021.
Helfrich, M., Dechow, R., Merl, S., Fuß, R., Räbiger, T., Kühling, I., Schlathölter, M., Kage, H., and Flessa, H.: Winter cover crops decreased soil mineral N contents and increased soil organic C stocks and N2O emission, Agr. Ecosyst. Environ., 367, 108985, https://doi.org/10.1016/j.agee.2024.108985, 2024.
Hillel, D.: Introduction to Environmental Soil Physics, 1st Edn., Academic Press, San Diego, CA, 511 pp., ISBN 9780123486554, 2003.
Intergovernmental Panel on Climate Change (IPCC): Climate Change 2022 – Mitigation of Climate Change: Working Group III Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge, https://doi.org/10.1017/9781009157926, 2022.
IPCC: 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories, Volume 4: Agriculture, Forestry and Other Land Use, edited by: Calvo Buendia, E., Tanabe, K., Kranjc, A., Baasansuren, J., Fukuda, M., Ngarize, S., Osako, A., Pyrozhenko, Y., Shermanau, P., and Federici, S., IPCC, Switzerland, ISBN 978-4-88788-232-4, 2019.
IUSS Working Group WRB: World Reference Base for Soil Resources 2014, update 2015: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, World Soil Resources Reports No. 106, FAO, Rome, ISBN 978-92-5-108369-7 (print), E-ISBN 978-92-5-108370-3 (PDF), 2015.
Jackson, R. B., Canadell, J., Ehleringer, J. R., Mooney, H. A., Sala, O. E., and Schulze, E. D.: A global analysis of root distributions for terrestrial biomes, Oecologia, 108, 389–411, https://doi.org/10.1007/BF00333714, 1996.
Jacobs, A., Poeplau, C., Weiser, C., Fahrion-Nitschke, A., and Don, A.: Exports and inputs of organic carbon on agricultural soils in Germany, Nutr. Cycl. Agroecosys., 118, 249–271, https://doi.org/10.1007/s10705-020-10087-5, 2020.
Jian, J., Du, X., Reiter, M. S., and Stewart, R. D.: A meta-analysis of global cropland soil carbon changes due to cover cropping, Soil Biol. Biochem., 143, 107735, https://doi.org/10.1016/j.soilbio.2020.107735, 2020.
Koch, H.-J., Grunwald, D., Essich, L., and Ruser, R.: Temporal dynamics of sugar beet (Beta vulgaris L.) N supply from cover crops differing in biomass quantity and composition, Front. Plant. Sci., 13, 920531, https://doi.org/10.3389/fpls.2022.920531, 2022.
Kravchenko, A. N., Toosi, E. R., Guber, A. K., Ostrom, N. E., Yu, J., Azeem, K., Rivers, M. L., and Robertson, G. P.: Hotspots of soil N2O emission enhanced through water absorption by plant residue, Nat. Geosci., 10, 496–500, https://doi.org/10.1038/ngeo2963, 2017.
Kühling, I., Mikuszies, P., Helfrich, M., Flessa, H., Schlathölter, M., Sieling, K., and Kage, H.: Effects of winter cover crops from different functional groups on soil-plant nitrogen dynamics and silage maize yield, Eur. J. Agron., 148, 126878, https://doi.org/10.1016/j.eja.2023.126878, 2023.
Lapierre, J., Machado, P. V. F., Debruyn, Z., Brown, S. E., Jordan, S., Berg, A., Biswas, A., Henry, H. A. L., and Wagner-Riddle, C.: Cover crop mixtures: A powerful strategy to reduce post-harvest surplus of soil nitrate and leaching, Agr. Ecosyst. Environ., 325, 107750, https://doi.org/10.1016/j.agee.2021.107750, 2022.
Lemke, R. L., Izaurralde, R. C., Malhi, S. S., Arshad, M. A., and Nyborg, M.: Nitrous Oxide Emissions from Agricultural Soils of the Boreal and Parkland Regions of Alberta, Soil Sci. Soc. Am. J., 62, 1096–1102, https://doi.org/10.2136/sssaj1998.03615995006200040034x, 1998.
Lenth, R. V., Bolker, B., Buerkner, P., Giné-Vázquez, I., Herve, M., Jung, M., Love, J., Miguez, F., Riebl, H., and Singmann, H.: emmeans: Estimated Marginal Means, aka Least-Squares Means, CRAN [code], https://rvlenth.github.io/emmeans/ (last access: 4 March 2024), 2025.
Li, X., Hu, F., and Shi, W.: Plant material addition affects soil nitrous oxide production differently between aerobic and oxygen-limited conditions, Appl. Soil Ecol., 64, 91–98, https://doi.org/10.1016/j.apsoil.2012.10.003, 2013.
Li, X., Petersen, S. O., Sørensen, P., and Olesen, J. E.: Effects of contrasting catch crops on nitrogen availability and nitrous oxide emissions in an organic cropping system, Agr. Ecosyst. Environ., 199, 382–393, https://doi.org/10.1016/j.agee.2014.10.016, 2015.
Lynch, M. J., Mulvaney, M. J., Hodges, S. C., Thompson, T. L., and Thomason, W. E.: Decomposition, nitrogen and carbon mineralization from food and cover crop residues in the central plateau of Haiti, SpringerPlus, 5, 973, https://doi.org/10.1186/s40064-016-2651-1, 2016.
Malnou, C. S., Jaggard, K. W., and Sparkes, D. L.: A canopy approach to nitrogen fertilizer recommendations for the sugar beet crop, Eur. J. Agron., 25, 254–263, https://doi.org/10.1016/j.eja.2006.06.002, 2006.
Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S. L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M. I., Huang, M., Leitzell, K., Lonnoy, E., Matthews, J. B. R., Maycock, T. K., Waterfield, T., Yelekçi, Ö., Yu, R., and Zhou, B. (Eds.): Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, https://doi.org/10.1017/9781009157896, 2021.
Millar, N. and Baggs, E. M.: Chemical composition, or quality, of agroforestry residues influences N2O emissions after their addition to soil, Soil Biol. Biochem., 36, 935–943, https://doi.org/10.1016/j.soilbio.2004.02.008, 2004.
Mitchell, J. P., Shrestha, A., and Irmak, S.: Trade-offs between winter cover crop production and soil water depletion in the San Joaquin Valley, California, J. Soil Water Conserv., 70, 430–440, https://doi.org/10.2489/jswc.70.6.430, 2015.
Muhammad, I., Sainju, U. M., Zhao, F., Khan, A., Ghimire, R., Fu, X., and Wang, J.: Regulation of soil CO2 and N2O emissions by cover crops: A meta-analysis, Soil Till. Res., 192, 103–112, https://doi.org/10.1016/j.still.2019.04.020, 2019.
Mutegi, J. K., Munkholm, L. J., Petersen, B. M., Hansen, E. M., and Petersen, S. O.: Nitrous oxide emissions and controls as influenced by tillage and crop residue management strategy, Soil Biol. Biochem., 42, 1701–1711, https://doi.org/10.1016/j.soilbio.2010.06.004, 2010.
Nasser, V.: Managing Soil Nitrogen Surplus: The Role of Winter Cover Crops in N2O Emissions and Carbon Sequestration, GRO.data, V1 [data set], https://doi.org/10.25625/HFEDA7, 2024.
Nouri, A., Lukas, S., Singh, S., Singh, S., and Machado, S.: When do cover crops reduce nitrate leaching? A global meta-analysis, Glob. Change Biol., 28, 4736–4749, https://doi.org/10.1111/gcb.16269, 2022.
Pan, F. F., Tang, J., and Chen, B. H.: Cover Crop Effects on Soil N Retention and Supply in Fertilizer-Intensive Cropping Systems (A Review), Eurasian Soil Sci.+, 55, 1278–1294, https://doi.org/10.1134/S1064229322090125, 2022.
Parkin, T. B., Kaspar, T. C., Jaynes, D. B., and Moorman, T. B.: Rye Cover Crop Effects on Direct and Indirect Nitrous Oxide Emissions, Soil Sci. Soc. Am. J., 80, 1551–1559, https://doi.org/10.2136/sssaj2016.04.0120, 2016.
Parr, M., Grossman, J. M., Reberg-Horton, S. C., Brinton, C., and Crozier, C.: Nitrogen Delivery from Legume Cover Crops in No-Till Organic Corn Production, Agron. J., 103, 1578–1590, https://doi.org/10.2134/agronj2011.0007, 2011.
Pelster, D. E., Chantigny, M. H., Rochette, P., Angers, D. A., Rieux, C., and Vanasse, A.: Nitrous Oxide Emissions Respond Differently to Mineral and Organic Nitrogen Sources in Contrasting Soil Types, J. Environ. Qual., 41, 427–435, https://doi.org/10.2134/jeq2011.0261, 2012.
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., Heisterkamp, S., Van Willigen, B., Ranke, J., and R Core Team: nlme: Linear and Nonlinear Mixed Effects Models, CRAN [code], https://svn.r-project.org/R-packages/trunk/nlme/ (last access: 4 March 2024), 2023.
Poeplau, C. and Don, A.: Carbon sequestration in agricultural soils via cultivation of cover crops – A meta-analysis, Agr. Ecosyst. Environ., 200, 33–41, https://doi.org/10.1016/j.agee.2014.10.024, 2015.
Qi, Z., Helmers, M. J., Malone, R. W., and Thorp, K. R.: Simulating Long-Term Impacts of Winter Rye Cover Crop on Hydrologic Cycling and Nitrogen Dynamics for a Corn-Soybean Crop System, T. ASABE, 54, 1575–1588, https://doi.org/10.13031/2013.39836, 2011.
R Core Team: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org/ (last access: 4 March 2024), 2023.
Ramirez-Garcia, J., Gabriel, J. L., Alonso-Ayuso, M., and Quemada, M.: Quantitative characterization of five cover crop species, J. Agric. Sci., 153, 1174–1185, https://doi.org/10.1017/S0021859614000811, 2015.
Riggers, C., Poeplau, C., Don, A., Bamminger, C., Höper, H., and Dechow, R.: Multi-model ensemble improved the prediction of trends in soil organic carbon stocks in German croplands, Geoderma, 345, 17–30, https://doi.org/10.1016/j.geoderma.2019.03.014, 2019.
Rochette, P., Angers, D. A., Chantigny, M. H., and Bertrand, N.: Nitrous Oxide Emissions Respond Differently to No-Till in a Loam and a Heavy Clay Soil, Soil Sci. Soc. Am. J., 72, 1363–1369, https://doi.org/10.2136/sssaj2007.0371, 2008.
Rösemann, C., Haenel, H.-D., Vos, C., Dämmgen, U., Döring, U., Wulf, S., Eurich-Menden, B., Freibauer, A., Döhler, H., Schreiner, C., Osterburg, B., and Fuß, R.: Calculations of gaseous and particulate emissions from German agriculture 1990–2019: Input data and emission results, Thünen Institute [data set], https://doi.org/10.3220/DATA20210323134503, 2021.
Rummel, P. S., Well, R., Pfeiffer, B., Dittert, K., Floßmann, S., and Pausch, J.: Nitrate uptake and carbon exudation – do plant roots stimulate or inhibit denitrification?, Plant Soil, 459, 217–233, https://doi.org/10.1007/s11104-020-04750-7, 2021.
Seitz, D., Fischer, L. M., Dechow, R., Wiesmeier, M., and Don, A.: The potential of cover crops to increase soil organic carbon storage in German croplands, Plant Soil, 488, 157–173, https://doi.org/10.1007/s11104-022-05438-w, 2023.
Sierra, C. A., Müller, M., and Trumbore, S. E.: Models of soil organic matter decomposition: the SoilR package, version 1.0, Geosci. Model Dev., 5, 1045–1060, https://doi.org/10.5194/gmd-5-1045-2012, 2012.
Signor, D. and Cerri, C. E. P.: Nitrous oxide emissions in agricultural soils: a review, Pesqui. Agropecu. Trop., 43, 322–338, https://doi.org/10.1590/S1983-40632013000300014, 2013.
Simmelsgaard, S. E.: The effect of crop, N-level, soil type and drainage on nitrate leaching from Danish soil, Soil Use Manage., 14, 30–36, https://doi.org/10.1111/j.1475-2743.1998.tb00607.x, 1998.
Smith, K. A., Ball, T., Dobbie, K. E., Massheder, J., and Rey, A.: Exchange of greenhouse gases between soil and atmosphere: interactions of soil physical factors and biological processes, Eur. J. Soil Sci., 54, 779–791, 2003.
Storr, T., Simmons, R. W., and Hannam, J. A.: Using frost-sensitive cover crops for timely nitrogen mineralization and soil moisture management, Soil Use Manage., 37, 427–435, https://doi.org/10.1111/sum.12619, 2021.
Taghizadeh-Toosi, A., Christensen, B. T., Hutchings, N. J., Vejlin, J., Kätterer, T., Glendining, M., and Olesen, J. E.: C-TOOL: A simple model for simulating whole-profile carbon storage in temperate agricultural soils, Ecol. Model., 292, 11–25, https://doi.org/10.1016/j.ecolmodel.2014.08.016, 2014.
Thangarajan, R., Bolan, N. S., Tian, G., Naidu, R., and Kunhikrishnan, A.: Role of organic amendment application on greenhouse gas emission from soil, Sci. Total Environ., 465, 72–96, https://doi.org/10.1016/j.scitotenv.2013.01.031, 2013.
Thapa, R., Mirsky, S. B., and Tully, K. L.: Cover Crops Reduce Nitrate Leaching in Agroecosystems: A Global Meta-Analysis, J. Environ. Qual., 47, 1400–1411, https://doi.org/10.2134/jeq2018.03.0107, 2018.
Tian, H., Xu, R., Canadell, J. G., Thompson, R. L., Winiwarter, W., Suntharalingam, P., Davidson, E. A., Ciais, P., Jackson, R. B., Janssens-Maenhout, G., Prather, M. J., Regnier, P., Pan, N., Pan, S., Peters, G. P., Shi, H., Tubiello, F. N., Zaehle, S., Zhou, F., Arneth, A., Battaglia, G., Berthet, S., Bopp, L., Bouwman, A. F., Buitenhuis, E. T., Chang, J., Chipperfield, M. P., Dangal, S. R. S., Dlugokencky, E., Elkins, J. W., Eyre, B. D., Fu, B., Hall, B., Ito, A., Joos, F., Krummel, P. B., Landolfi, A., Laruelle, G. G., Lauerwald, R., Li, W., Lienert, S., Maavara, T., MacLeod, M., Millet, D. B., Olin, S., Patra, P. K., Prinn, R. G., Raymond, P. A., Ruiz, D. J., van der Werf, G. R., Vuichard, N., Wang, J., Weiss, R. F., Wells, K. C., Wilson, C., Yang, J., and Yao, Y.: A comprehensive quantification of global nitrous oxide sources and sinks, Nature, 586, 248–256, https://doi.org/10.1038/s41586-020-2780-0, 2020.
Tonitto, C., David, M. B., and Drinkwater, L. E.: Replacing bare fallows with cover crops in fertilizer-intensive cropping systems: A meta-analysis of crop yield and N dynamics, Agr. Ecosyst. Environ., 112, 58–72, https://doi.org/10.1016/j.agee.2005.07.003, 2006.
Unger, P. W. and Vigil, M. F.: Cover crop effects on soil water relationships, J. Soil Water Conserv., 53, 200–207, 1998.
VDLUFA: Bestimmung von mineralischem Stickstoff (Nitrat und Ammonium) in Bodenprofilen (NminLabormethode A 6.1.4.1), 2002.
Voisin, A.-S., Salon, C., Munier-Jolain, N. G., and Ney, B.: Quantitative effects of soil nitrate, growth potential and phenology on symbiotic nitrogen fixation of pea (Pisum sativum L.), Plant Soil, 243, 31–42, https://doi.org/10.1023/A:1019966207970, 2002.
Wagner-Riddle, C., Congreves, K. A., Abalos, D., Berg, A. A., Brown, S. E., Ambadan, J. T., Gao, X., and Tenuta, M.: Globally important nitrous oxide emissions from croplands induced by freeze–thaw cycles, Nat. Geosci., 10, 279–283, https://doi.org/10.1038/ngeo2907, 2017.
Wang, J., Zhang, S., Sainju, U. M., Ghimire, R., and Zhao, F.: A meta-analysis on cover crop impact on soil water storage, succeeding crop yield, and water-use efficiency, Agr. Water Manage., 256, 107085, https://doi.org/10.1016/j.agwat.2021.107085, 2021.
Wayman, S., Cogger, C., Benedict, C., Burke, I., Collins, D., and Bary, A.: The influence of cover crop variety, termination timing and termination method on mulch, weed cover and soil nitrate in reduced-tillage organic systems, Renew. Agr. Food Syst., 30, 450–460, https://doi.org/10.1017/S1742170514000246, 2015.
Weitz, A. M., Linder, E., Frolking, S., Crill, P. M., and Keller, M.: N2O emissions from humid tropical agricultural soils: effects of soil moisture, texture and nitrogen availability, Soil Biol. Biochem., 33, 1077–1093, https://doi.org/10.1016/S0038-0717(01)00013-X, 2001.
Wickham, H., Pedersen, T. L., Seidel, D., and Thomas, L. P.: scales: Scale Functions for Visualization, CRAN [code], https://scales.r-lib.org (last access: 4 March 2024), 2023.
Zuur, A. F., Ieno, E. N., Walker, N., Saveliev, A. A., and Smith, G. M.: Mixed effects models and extensions in ecology with R, Springer New York, NY, https://doi.org/10.1007/978-0-387-87458-6, 2009.